|
|
|
Copyright © 2004 by the author(s). Published here under license by The Resilience Alliance.
Go to the pdf version of this article.
The following is the established format for referencing this article:
Wikelski, M., J. Foufopoulos, H. Vargas, and H. Snell. 2004. Galápagos Birds and Diseases: Invasive Pathogens as Threats for Island Species. Ecology and Society 9(1): 5. [online] URL: http://www.ecologyandsociety.org/vol9/iss1/art5/
A version of this article in which text, figures, tables, and appendices are separate files may be found by following this link.
Insight Galápagos Birds and Diseases: Invasive Pathogens as Threats for Island Species Martin Wikelski1, Johannes Foufopoulos, Hernan Vargas, and Howard Snell2
1Princeton University; 2Department of Biology, University of New Mexico / Charles Darwin Foundation
Exotic diseases and parasites have caused extinctions on islands and
continents, particularly when they spread through assemblages of
immunologically naïve species. Hawaii has lost a substantial part of its
endemic bird fauna since the introduction of avian malaria at the beginning of
the 20th century. In contrast, the Galápagos archipelago still possesses its
entire endemic avifauna. Several of these Galápagos bird populations are in decline,
however, and wildlife managers seek guidance to counteract a potential man-made
ecological disaster. We recommend that endemic birds be tested for
susceptibility to disease outside the Galápagos so that protection efforts can be
better designed to deal with actual threats. At present, the best and perhaps
only management option is to protect the isolation of these island communities
because treating or vaccinating wild bird populations against diseases is almost
impossible. If the isolation of the Galápagos Islands is successful, we will
preserve the complete avifauna of an archipelago for the first time in the
history of human colonization in the Pacific eco-region.
KEY WORDS: Endemic birds, Galápagos, extinctions, introduced diseases, isolation, susceptible species, wildlife management.
Published: January 21, 2004
INTRODUCTION
One of the most important factors contributing to extinctions on islands is the introduction of exotic organisms (Diamond 1984, Groombridge 1992, Dobson and Foufopoulus 2001). Among exotic organisms, pathogens have been largely ignored by conservation biologists until recently (Altizer et al. 2001). This can be attributed in part to the cryptic and frequently transient nature of parasite infection and in part to the difficulty of collecting disease data in the field. A careful review of the literature indicates that pathogenic organisms have been involved in numerous declines and extinctions of endemic species on oceanic and land-bridge islands (Diamond 1994; Table 1).
|
Table 1. Examples of disease outbreaks with conservation implications on islands.
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
DISEASE SUSCEPTIBILITY AND EXTINCTION
IN ISLAND
POPULATIONS
The Galápagos Islands have not experienced recent extirpations of bird species, although one exception may be the demise of the Floreana Mockingbird (Nesomimus trifasciatus) which is now found only on Floreana’s satellite island, Champion (D. J. Anderson, personal communication). This situation is remarkable given that island populations are highly susceptible to extinction (Groombridge 1992). In other oceanic archipelagos, many bird species were extirpated soon after prehistoric humans reached them. For example, more than 200 species of extinct island birds are only recorded as only subfossils on islands in the Pacific Ocean. These probably vanished following settlement of the islands by Polynesians during the Austronesian expansion (Milberg and Tyrberg 1993). In addition, several species now considered endemic to single islands or island groups had a much wider distribution in the past (Smith et al. 1995, Steadman 1995). The anthropogenic mechanisms resulting in the demise of island taxa are complex and include habitat destruction, predation by humans, and the introduction of exotic predators, competitors, and pathogens (Diamond and Veitch 1981, Holmes 1996). Although most, if not all, of these factors also affect mainland species, the small size and naïve nature of most island populations render them particularly prone to extinction (Atkinson et al. 1995, Cole et al. 1995).
A well-documented example of indigenous island hosts adversely affected by exotic pathogens is the introduction of avian poxvirus and avian malaria, Plasmodium relictum, into the avifauna of the Hawaiian Islands (Warner 1968, Van Riper et al. 1986, Jenkins et al. 1989, Atkinson et al. 1995, Atkinson et al. 2000, Work et al. 2000). Similar but less well-documented examples exist from Christmas Island in the Indian Ocean where the extinction of two endemic species of rodent appear to be linked to the introduction of a trypanosome pathogen (Day 1981, Pickering and Norris 1996, see also Fancy and Snetsinger 2001). Epidemics that have devastated island taxa have also been reported for lacertid lizards in the Tyrrhenian Sea and for birds along the eastern seaboard of the United States (Day 1981).
Endemic island species held in captivity appear similarly sensitive to infection by exotic pathogens. A captive breeding program for the Pink Pigeon (Nesoenas mayeri), an endangered species from Mauritius, encountered serious problems after the hatchlings were infected with a pigeon herpesvirus from asymptomatic Rock Doves (Columba livia) used as foster parents (Snyder et al. 1985). Another Mascarene endemic, the Mauritius Kestrel (Falco punctatus) was shown to be sensitive to herpesvirus hepatitis (Cooper et al. 1981). Penguins, which are not island endemics per se but have evolved mainly in the absence of diverse pathogen communities, have been shown to be very susceptible to infectious pathogens, most notably various Plasmodium species (Graczyk et al. 1995).
Several mechanisms are blamed for the predisposition of island populations to exotic pathogens. Both theoretical and empirical evidence suggests that small host populations on isolated islands support only impoverished parasite communities (Dobson et al. 1992, Dobson and McCallum 1997). This is because resident parasites are subject to the same constraints that limit host diversity. They are also subject to additional demographic and genetic constraints dictated by the small population sizes of their hosts. As a result, island endemics have been exposed to few parasites and other pathogens during their recent evolutionary history. Long-term release from parasitism could result in a partial loss of energetically expensive immune systems in isolated island populations (K. Lindström, personal communication). Host immune responses could be particularly compromised by the loss of genetic diversity in small, isolated populations (Lyles and Dobson 1993). Within inbred island populations, loss of heterozygosity has been shown to result in reduced major histocompatibility complex diversity and also in subsequent selection against inbred individuals (Keller et al. 1994).
In summary, the evolution of island taxa in a parasite-scarce environment, alone or in combination with a long-term loss of genetic diversity, often results in a compromised ability to respond to new pathogens.
CURRENT AVIAN DISEASE PROBLEMS
ON THE GALAPAGOS
Despite isolation and the short duration of human settlement on the Galápagos Islands, several exotic pathogens have been introduced to the archipelago and appear to be affecting wildlife populations. However, no hard evidence has been gathered to support this statement unambiguously.
The introduction of some of these parasites may be related to the importation and maintenance of domestic chickens which share the habitat of endemic birds on islands inhabited by humans. Evidence for or against disease transmission via chickens is so far missing; it is currently being studied (P. Parker, personal communication). During periods of abundant food, supply chickens may become feral and invade National Park areas, a circumstance documented during the El Niño year of 1988 (Vargas 1999). This increases the risk of disease transmission to indigenous birds.
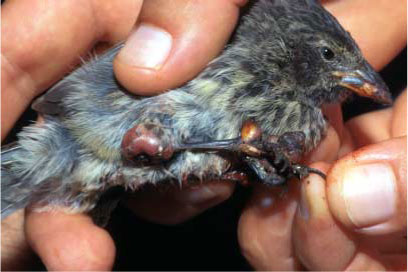
Infectious diseases, such as Newcastle disease, Marek’s disease, avian
mycoplasmosis and avian pox (Fig. 1), have been recorded
in domestic chickens farmed on the islands (Vargas and Snell 1997). Avian pox
now regularly occurs in several species of Darwin’s finches (Fig.
2); Galápagos Mockingbirds, Nesomimus parvulus (Vargas 1987); and
Yellow Warblers, Dendroica petechia (Duffy and Harcourt 1981).
This disease may have been brought to the island during the first human visits.
As early as 1905, the journal of the California Academy of Sciences expedition
mentions lesions that could be interpreted as avian pox (E.W. Gifford, unpublished manuscript; B. West, personal
communication). Several known vectors of diseases have been more recently
introduced, further increasing the risk of disease transmission. For example,
the mosquito Culex quinquefasciatus, a vector of avian malaria (Plasmodium
spp.), was recently introduced to the Galápagos (H. Vargas, personal
observation). It is unclear whether avian malaria already exists on the
islands; it has not been detected. Bird surveys conducted in the past four
years indicate that several of the island’s bird species have declined in
population and range. Diseases are suspected among the reasons for these
declines (H. Vargas, personal observation). Most of the declines have
taken place on inhabited islands where introduced bird species are most
abundant. One of the most endangered birds now appears to be the San Cristóbal
Vermilion Flycatcher (Pyrocephalus rubinus dubius). Not a single
individual of this species was observed during a six-month survey in 1998
(Vargas and Bensted-Smith 2000). A long-term, steady decline in population size
is clearly distinct from the frequent, normal population fluctuations of native
bird fauna on the Galápagos Islands (cf. Grant and Grant 1980, 1993). On the
other hand, some potentially sensitive species have avoided some of these
pathogens. For example, Galápagos Penguins (Spheniscus mendiculus) appear free of
avian malaria and Marek’s disease (Miller et al. 2001). However, penguins may
simply be less susceptible to mosquito-transmitted diseases because they inhabit
coastal habitats far from mosquitoes.
|
Fig. 1. A Darwin’s finch
affected by avian pox (St. Cruz Island, Galápagos, 2000).
|
|
Fig. 2. A domestic
chicken with avian pox (St. Cruz Island, Galápagos, 2000).
|

Rock Doves introduced to the Galápagos have carried with them Trichomonas gallinae. This parasite has now been recorded in the endemic Galápagos Dove, Zenaida galapagoensis (Harmon et al. 1987). The impact of this pathogen on the endemic dove species is not known. Nevertheless, populations of Galápagos Dove appear to be rapidly declining on inhabited islands. A new botfly species (Philornis downsi ), a parasite known to significantly lower fledgling success, was recently detected on nestlings of several species of Darwin’s finches, Galápagos Mockingbirds, Yellow Warblers and the Dark-billed Cuckoo, Coccyzus melacoryphus (Fessl and Tebbich 2002). Parasitic nematodes (Dispharynx spiralis ) were found in the endemic Dark-billed Cuckoos and may have been partially responsible for the high cuckoo mortality observed in 1998 (Vargas and Bensted-Smith 2000). It is not known whether these parasites are endemic to the Galápagos or were introduced from feral animals.
POTENTIAL AVIAN DISEASE THREATS
TO THE GALÁPAGOS
Table 2 provides a comprehensive list of the most important potential disease threats to Galápagos birds identified by an interdisciplinary group of biologists (International Workshop on Avian Diseases 2000). Although most of the pathogens listed have caused conservation problems in other regions of the world, it is difficult to predict the extent to which they could affect Galápagos bird populations; the effect of an introduced pathogen is dependent upon the intricate interplay between host, parasite and prevailing environmental conditions (Aron and Patz 2000). For example, although avian malaria is a serious conservation problem on the Hawaiian Islands, this disease may not spread through certain populations of Galápagos birds because the requisite mosquito vectors are not well suited for the relatively arid environment. However, environmental conditions could provide opportunities for the successful spread of malaria and other diseases if these pathogens reach the humid Galápagos highlands or if coastal parts of the islands become wetter because of climate change, e.g., increased frequency and intensity of El Niño.
|
Table 2. Diseases that
cause major mortality in wild birds, methods of transmission, and methods of
control.
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Another potential threat is West Nile encephalitis, a serious avian disease that spread rapidly across North America after its introduction to New York City in 1999 (Lanciotti et al. 1999). Cases of this viral disease have now been documented on the western coast of the United States (Causey et al. 2003, Rappole and Hubalek 2003). The virus is expected to be transported to Central and South America, and from there to the Galápagos by migrating shore and sea birds (cf. Rappole et al. 2000, Malakoff 2002, Malkinson et al. 2001, 2002, Zeller and Murgue 2001, Causey et al. 2003). So far, it is unknown how island birds will react to this virus, and whether the virus will remain as destructive as it is currently.
Most of the other pathogens listed in Table 2 are widespread, opportunistic, bacterial, or viral microparasites. In general, they infect a wide range of hosts. If these pathogens become established in Galápagos bird populations, they have the potential to cause widespread die-offs. Although these diseases have not yet been implicated in any global species extinctions, their introduction to a small isolated ecosystem like that of the Galápagos archipelago could have catastrophic results for the local bird taxa.
WHAT CAN WE DO TO PREVENT
AN ECOLOGICAL DISASTER?
Past experience from continental North America indicates that, once an exotic pathogen becomes established in a wildlife population, few, if any, effective mechanisms exist to control its transmission. Although the spread and/or persistence of a disease on an island may be somewhat different from its spread and/or persistence on the mainland, the principles of disease establishment are likely to prove similar in both scenarios. The culling of infected individuals from natural populations may help in disease control when host densities are high (Ferguson et al. 2001), such as at breeding farms or densely visited stopover habitats (Rocke and Brand 1994). Vector control is often the method of choice to stop the transmission of mosquito-borne diseases (Hougard et al. 2002). Bird behavior can be influenced as well so that pathogen transmission becomes less likely. For example, influence can result in the exclusion of infected birds from conspecific flocks. However, exclusion of birds may work only in very small and specific locations, such as garbage dumps, and may not be effective in natural areas. The transmission of specific diseases like botulism (Rocke and Brand 1994) can be achieved by wildlife managers who simply pick up infectious carcasses, change the environment, e.g., burn habitat, or eliminate pathogen reservoir species. However, in all cases considered by wildlife experts so far, the possibilities for reducing disease transmission are very limited. This will be particularly true in the Galápagos Islands. The main reason for these management limits in the Galápagos are that many areas of the archipelago are difficult and expensive to reach. Conservation managers also have to keep in mind the political realities of an economy based on eco-tourism. Extreme measures, such as the culling of birds, are hard to prescribe when eco-tourists want to see those same birds, even if rare.
In light of the inability to control a disease epidemic once it has been established on the Galápagos Islands, emphasis needs to be placed on preventing the introduction of pathogens to the archipelago. Unfortunately, options to protect Galápagos birds against the introduction of diseases are limited. First, authorities should protect and guard the ecological isolation of the islands via activities supplemental to the existing Galápagos quarantine program (Trillmich 1992). The current quarantine program attempts to prevent the introduction of foreign organisms by surveying shipments at Ecuadorian ports before organisms can reach the Galápagos archipelago. Additional activities should emphasize control for alien vectors of avian diseases and parasites. For example, the number of chickens on the islands should be reduced to the extent possible, even if there have not been any obvious disease transmissions documented between domestic chickens and wild birds.
The second necessary activity to help prevent the further spread of disease is a realistic baseline monitoring program to allow early detection of novel avian diseases or parasites in the Galápagos. This means a modest veterinary pathology laboratory should be established and operated. A list of opportunistic activities should also be set out to ensure that collaborating scientists and managers work to increase monitoring efficacy. Fortunately, both programs have already been started (H. Snell, personal communication).
Third, conservation ecologists and managers need detailed contingency plans in place to respond to cases of abnormal morbidity or mortality. These plans could include provisions for the rapid export of samples from infected animals to international centers for disease control and detection. A contingency plan should also include an evaluation of the intervention options available for those pathogens most likely to threaten native species. It is imperative to test, outside of the Galápagos, whether and to what degree land birds, such as Darwin’s finches, are susceptible to several major avian diseases like malaria or the West Nile virus. If Darwin’s finches are indeed susceptible to these diseases, a long-term captive breeding program at a disease-free location should be considered to prevent and anticipate problems of captive propagation before populations decline too far (cf. Jenkins et al. 1989). Information on disease susceptibility would also help to strengthen quarantine procedures and allow for more political leeway in conservation efforts.
Of the limited options for disease control currently available to the Galápagos Islands, vector control through education and eradication programs around human settlements may be the most promising. Such practices, although seemingly limited, can go a long way toward preventing the introduction of pathogens exotic to the Galápagos Islands or reducing their spread. Resources should be committed to building diagnostic laboratories, collecting baseline data, and training personnel.
THE FUTURE
The Galápagos Islands provide a rare chance to preserve the intact endemic avian communities of an archipelago because no species of Galápagos birds has yet gone extinct. A powerful legal framework exists to support the activities necessary to reduce the arrival of new diseases, and global concern over the loss of biological diversity in the area is increasing. The islands therefore represent a unique opportunity to successfully apply the lessons in wildlife disease control and prevention that have been learned in other areas of the world (Friend et al. 2001). Because some populations of Galápagos birds are in decline, local management agencies are seeking guidance for the formulation of effective regulations. The opportunity to conserve a natural laboratory of evolution as a showcase for humanity could be lost quickly unless global concern is translated into effective and appropriate management (Loope et al. 2001).
Responses to this article are invited. If accepted for publication, your response will be hyperlinked to the article. To submit a comment, follow this link. To read comments already accepted, follow this link.
Acknowledgments
We thank all participants of the International Workshop on Avian Diseases
held at Princeton University in 2000. Victor Apanius, Charlotte
Causton, Andy Dobson, Birgit Fessl, Joe Flanagan, Peter Grant, Rosemary Grant,
Michaela Hau, Philippe Heeb, Nigella Hillgarth, Bruce Victor Hofkin, Andrea
Gager, Susan Jarvi, Richard Julian, William Karesh, Uriel Kitron, Dennis
LaPointe, Robert McLean, Carlos Moreno, Robert Ricklefs, Washington Tapia, and
Thierry Work all contributed to this project. We also thank Barbara West for
analyzing historical records. We are grateful for the workshop funding provided by the Charles Darwin Foundation, Inc., and the Swiss Association of Friends of the Galápagos Islands, as well as the University of Illinois and Princeton University.
Altizer, S., J. Foufopoulos, and A. Gager. 2001. Conservation and diseases. Pages 109-126 in S. Levin, editor. Encyclopedia of biodiversity. Academic Press, San Diego, California, USA.
Aron, J. L., and J. Patz. 2000. Ecosystem change and public health: A global perspective. Johns Hopkins University Press, Baltimore, USA.
Atkinson, C. T., K. L. Woods, R. J. Dusek, L. S. Sileo, and W. M. Iko. 1995. Wildlife disease and conservation in Hawaii—Pathogenicity of avian malaria (Plasmodium relictum) in experimentally infected Iiwi (Vestiaria coccinea). Parasitology 111:S59-S69.
Atkinson, C. T., R. J. Dusek, K. L. Woods, and W. M. Iko. 2000. Pathogenicity of avian malaria in experimentally-infected Hawaii Amakihi. Journal of Wildlife Diseases 36:197-204.
Brasil, M. 1910. Un oiseau éteint de la Reunion, Fregilupus varius (Bodd.). Bulletin Société Linne Normandie 6:1-29.
Causey, D., J. Trimble, W. Hallwachs, D. Brooks, and D. Janzen. 2003. Migratory birds and the spread of West Nile. Science 299:821.
Cooper, J. E. 1993. Historical survey of disease in birds. Journal of Zoo and Wildlife Medicine. 24:256-264.
Cole F. R., L. L. Loope, A. C. Medeiros, J. A. Raikes and C. S. Wood. 1995. Conservation implications of introduced game birds in high-elevation Hawaiian shrubland. Conservation Biology 9:306-313.
Cooper, J. E., C. G. Jones, and A. W. Owadally. 1981. Morbidity and mortality in the Mauritius Kestrel (Falco punctatus). Pages 31-35 in J. E. Cooper and A. G. Greenwood, editors. Recent advances in the study of raptor diseases. Chiron, Keighley, UK.
Day, D. 1981. The doomsday book of animals. Viking, New York, USA.
Diamond, J. 1984. Historic extinctions: a Rosetta stone for understanding prehistoric extinctions. Pages 824-862 in P. S. Martin and R. G. Klein, editors. Quaternary extinctions: a prehistoric revolution. University of Arizona Press, Tucson, Arizona, USA.
Diamond, J. 1994. Ecological collapses of past civilizations. Proceedings of the American Philosophical Society 138:363-370
Diamond, J., and C. R. Veitch. 1981. Extinctions and introductions in the New Zealand avifauna: cause and effect? Science 211:499-501.
Dobson, A. P., S. V. Pacala, J. D. Roughgarden, E. R. Carper, and E. A. Harris. 1992. The parasites of Anolis lizards in the northern Lesser Antilles. I. Patterns of distribution and abundance. Oecologia 91:110-117.
Dobson, A. P., and H. McCallum. 1997. The role of parasites in bird conservation. Pages 155-173 in D. H. Clayton and J. Moore, editors. Host-parasite evolution: general principles and avian models. Oxford University Press, Oxford, UK.
Dobson, A. P., and J. Foufopoulos. 2001. Emerging infectious pathogens of wildlife. Philosophical Transaction of the Royal Society of London B 356:1001-1012.
Duffy, D., and S. Harcourt. 1981. An Investigation of Avian Pox Disease: Its Distribution and Effects on Galápagos Birds. Pages 108-111 in D. C. Duffy, D. J. Reynolds, and M. J. Campos, editors. 1980 Annual Report of the Charles Darwin Research Station. Charles Darwin Foundation, Galápagos, Ecuador.
Fancy, S. G., and T. J. Snetsinger. 2001. What caused the population decline of the bridled white-eye on Rota, Mariana Islands? Studies in Avian Biology 22:274-280.
Ferguson, N.M., C.A. Donnelly, and R. M. Anderson. 2001. Transmission intensity and impact of control policies on the foot and mouth epidemic in Great Britain. Nature 413:542-548.
Fessl, B., and S. Tebbich. 2002. Philornis downsi—a recently discovered parasite on the Galápagos archipelago—a threat for Darwin’s finches? Ibis 144:445–451.
Friend, M., R. G. McLean, and F. J. Dein. 2001. Disease emergence in birds: challenges for the twenty-first century. The Auk 118:290-303.
Gassmann-Duvall, R. 1987. An acute Cyathostoma bronchialis outbreak in the Hawaiian goose and other parasitic findings. Page 61-68 in Annual Proceedings of the American Association of Zoo Veterinarians. American Association of Zoo Veterinarians, Atlanta, Georgia, USA.
Graczyk, T. K., J. F. Cockrem, M. R. Cranfield, J. T. Darby, and P. Moore. 1995. Avian malaria seroprevalence in wild New Zealand penguins. Parasite 2:401-405.
Grant, P. R., and B. R. Grant. 1980. Annual variation in finch numbers, foraging and food supply on Isla Daphne Major, Galápagos. Oecologia 46:55-62.
Grant, B. R., and P. R. Grant. 1993. Evolution of Darwin's finches caused by a rare climatic event. Proceedings of the Royal Society of London B 251:111-117.
Groombridge, B. 1992. Global Biodiversity - Status of the Earth's Living Resources: A Report Compiled by the World Conservation Monitoring Centre. Chapman and Hall, London, New York.
Harmon, W. M., W. A. Clark, A. C. Hawbecker, and M. Stafford. 1987. Trichomonas gallinae in columbiform birds from the Galápagos Islands. Journal of Wildlife Diseases 23:492-494.
Holmes, J. C. 1996. Parasites as threats to biodiversity in shrinking ecosystems. Biodiversity and Conservation 5:975-983.
Hougard, J. M., D. Fontenille, F. Chandre, F. Darriet, P. Carnevale, and P. Guillet. 2002. Combating malaria vectors in Africa: current directions of research. Trends in Parasitology 18:283-286.
International Workshop on Avian Diseases. 2000. Princeton University, Princeton, New Jersey, USA.
Jenkins, C. D., S. A. Temple, C. Van Riper, and W. R. Hansen. 1989. Disease-related aspects of conserving the endangered Hawaiian crow. Pages 77-87 in J. E. Cooper, editor. Disease and threatened birds. ICBP Technical Publication Number 10. International Council for Bird Preservation, Cambridge, UK.
Keller, L. F., P. Arcese, J. N. M. Smith, W. M. Hochachka, and S. C. Stearns. 1994. Selection against inbred song sparrows during a natural population bottleneck. Nature 372:356-357.
Lanciotti, R. S., J. T. Roehrig, V. Deubel, J. Smith, M. Parker, K. Steele, B. Crise, K. E. Volpe, M. B. Crabtree, J. H. Scherret, R. A. Hall, J. S. MacKenzie, C. B. Cropp, B. Panigrahy, E. Ostlund, B. Schmitt, M. Malkinson, C. Banet, J. Weissman, N. Komar, H. M. Savage, W. Stone, T. McNamara, and D. J. Gubler. 1999. Origin of the West Nile virus responsible for an outbreak of encephalitis in the northeastern United States. Science 286:2333-2337.
Loope, L. L., F. G. Howarth, F. Kraus, and T. K. Pratt. 2001. Newly emergent and future threats of alien species to Pacific birds and ecosystems. Studies in Avian Biology 22:291-304.
Lyles, A. M., and A. P. Dobson. 1993. Infectious disease and intensive management: population dynamics, threatened hosts and their parasites. Journal of Zoo and Wildlife Medicine 24:315-326.
Malakoff, D. 2002. Infectious disease: Bird advocates fear that West Nile virus could silence the spring. Science 297:1989-1989.
Malkinson, M., C. Banet, Y. Weisman, S. Pokamonski, R. King, and V. Deubel. 2001. Intercontinental transmission of West Nile virus by migrating white storks. Emerging Infectious Diseases 7:540.
Malkinson, M., C. Banet, Y. Weisman, S. Pokamonski, R. King, M.T. Drouet, and V. Deubel. 2002. Introduction of West Nile virus in the Middle East by migrating white storks. Emerging Infectious Diseases 8:392-397.
McCallum, H., and A. Dobson. 1995. Detecting disease and parasite threats to endangered species and ecosystems. Trends in Ecology and Evolution 10:190-194.
Milberg, P., and T. Tyrberg. 1993. Naive birds and noble savages—a review of man-caused prehistoric extinctions of island birds. Ecography 16:229-250.
Miller, G. D., B. V. Hofkin, H. Snell, A. Hahn, and R. D. Miller. 2001. Avian malaria and Marek’s disease: potential threats to Galápagos penguins Sphiniscus mendiculus. Marine Ornithology in press.
Pickering J., and C. A. Norris. 1996. New evidence concerning the extinction of the endemic murid Rattus macleari from Christmas Island, Indian Ocean. Australian Mammalogy 19:19-25.
Ralph, C. J., and C. Van Riper. 1985. A historical perspective on the decline in native Hawaiian birds. Pages 7-42 in S. Temple, editor. Bird Conservation. University of Wisconsin Press, Madison, Wisconsin, USA.
Rappole, J. H., S. R. Derrickson, and Z. Hubalek. 2000. Migratory birds and spread of West Nile virus in the Western Hemisphere. Emerging Infectious Diseases 6:319-328.
Rappole, J. H., and Z. Hubalek. 2003. Migratory birds and West Nile virus. Journal of Applied Microbiology 94:47S-58S.
Rocke, T. E., and C. J. Brand. 1994. Use of sentinel mallards for epizootiologic studies of avian botulism. Journal of Wildlife Diseases 30:514-522.
Smith, T. B, L. A. Freed, J. K. Lepson, and J. H. Carothers. 1995. Evolutionary consequences of extinctions in populations of a Hawaiian honeycreeper. Conservation Biology 9:107-113.
Snyder B., J. Thilsted, B. Burgess, and M. Richard. 1985. Pigeon herpesvirus mortalities in foster reared Mauritius pink pigeons. Pages 69-70 in Annual Proceedings of the American Association of Zoo Veterinarians. American Association of Zoo Veterinarians, Atlanta, Georgia, USA.
Steadman, D. W. 1995. Prehistoric extinctions of Pacific island birds: biodiversity meets zooarchaeology. Science 267:1123-1131.
Trillmich, F. 1992. Conservation problems on Galápagos—the showcase of evolution in danger. Naturwissenschaften 79:1-6.
Van Riper, C., S. G. van Riper, M. L. Goff, and M. Laird. 1986. The epizootiology and ecological significance of malaria on the birds of Hawaii. Ecological Monographs 56:327-344.
Vargas, H. 1987. Frequency and effect of Pox-like Lesions in Galápagos Mockingbirds. Journal of Field Ornithology 58:101-102.
Vargas, H. 1999. Ornithological notes from Galápagos in 1998. Page 72 in P. Ospina and E. Muñoz, editors. Galápagos Report 1998-1999. World Wildlife Fund-Fundacion Natura, Quito, Ecuador.
Vargas, H, and H. L. Snell. 1997. The arrival of Marek’s disease to Galápagos. Noticias de Galápagos 58:4-5.
Vargas, H., and R. Bensted-Smith. 2000. Past and present ornithology in Galápagos. Pages 47-52 in N. Sitwell, L. Baert and G. Cuppois, editors. Proceedings of the Symposium Science and Conservation in Galápagos. Royal Belgian Institute of Natural Sciences, Brussels, Belgium.
Veterinary Record. 1992. Other birds. Veterinary Record 131:208
Warner, R. E. 1968. The role of introduced diseases in the extinction of the endemic Hawaiian avifauna. Condor 70:101-120.
Work, T.M., J.G. Massey, B.A. Rideout, C.H. Gardiner, D. Ledig, and J.P. Dubey. 2000. Toxiplasmosis in free-ranging Hawaiian crows (Corvus hawaiiensis). Journal of Wildlife Diseases 36:205-212.
Zeller, H. G., and B. Murgue. 2001. The role of migrating birds in the West Nile virus epidemiology. Medecine Et Maladies Infectieuses 31:168S-174S.
Address of Correspondent:
Martin Wikelski
Department of Ecology and Evolutionary Biology
Princeton University
Princeton, New Jersey 08544-1003 USA
Phone: (609) 258-6133
Fax: (609) 258-1682
wikelski@princeton.edu
![]()