|
|
|
Copyright © 2004 by the author(s). Published here under license by The Resilience Alliance.
Go to the pdf version of this article.
The following is the established format for referencing this article:
Martin, S. 2004. The cost of restoration as a way of defining resilience: a viability approach applied to a model of lake eutrophication. Ecology and Society 9(2): 8. [online] URL: http://www.ecologyandsociety.org/vol9/iss2/art8/
Report The Cost of Restoration as a Way of Defining Resilience: a Viability Approach Applied to a Model of Lake Eutrophication Sophie Martin
Cemagref
- Abstract
- Introduction
- Methods
- Results
- Discussion
- Speculation
- Responses to this Article
- Acknowledgments
- Literature Cited
Multiple stable states or alternative equilibria in ecological systems have been recognized since the 1960s in the ecological literature. Very often, the shift between alternative states occurs suddenly and the resource flows from these systems are modified. Resilience is the capacity of a system to undergo disturbance and maintain its functions and controls. It has multiple levels of meaning, from the metaphorical to the specific. However, most studies that explore resilience-related ideas have used resilience as a metaphor or theoretical construct. In a few cases, it has been defined operationally in the context of a model of a particular system. In this paper, resilience is defined consistently with the theoretical uses of the term, in the context of ecosystem models within an application to a simple model of lake eutrophication. The theoretical definitions of resilience and the characteristics of the operational definition that are necessary for ensuring consistency are reviewed. A mathematical formulation of resilience is built in the framework of the viability theory. This formulation emphasizes the link between resilience and the cost of the recovery after a disturbance. This cost is first chosen in relation to the time of crisis in the application to a model of lake eutrophication. The resilience values are then obtained by numerical integration. For another choice of the cost function, the viability algorithm is needed to compute the resilience values. These applications demonstrate the usefulness of our operational definition.
KEY WORDS: dynamic systems, ecosystem models, eutrophication, lake ecosystem, resilience, time of crisis.
Published: October 15, 2004
Over recent decades, humans have imposed greater and greater demands upon natural systems. Some natural resources, such as fossil fuel, are in fact essential to modern society; other natural resources, such as air and water, are essential to human life. Consequently, the sustainability of the resource flows from these natural systems is a crucial issue that many researchers have already raised (Odum 1993, Arrow et al. 1995).
The term “resilience” is often used when the sustainability of a system is studied. It has multiple levels of meaning, from the metaphorical to the specific (Carpenter et al. 2001). In the context of ecosystem modeling, the purpose of this paper is to provide an operational dynamic definition of resilience, consistent with the metaphorical and theoretical uses of the term. Using the framework of the viability theory, we first propose a mathematical formulation of resilience that is consistent with the characteristics taken from the theoretical definitions. Then, we apply this definition of resilience to a simple model of lake eutrophication to demonstrate the usefulness of our operational definition with regard to the viability algorithm. As part of the rationale for the methods described here, the different conceptual and operational definitions of resilience are briefly compared and reviewed because our operational definition must be consistent with the conceptual one.
A number of conceptual definitions of resilience have been proposed. For Pimm (1984), resilience is the ability of a system to resist disturbance and the rate at which it returns to equilibrium following a disturbance. Loss of resilience is due to slow dynamics near a stable equilibrium. This definition only applies to the behavior of a linear system or a nonlinear one in the immediate vicinity of a stable equilibrium where a linear approximation is valid (Pimm 1991). As for Gunderson and Holling (2002), resilience is the capacity of a system to undergo disturbance and maintain its functions and controls. Loss of resilience is associated with slow dynamics in a region that separates domains of attraction. This definition refers to the behavior of a nonlinear system near the boundary of a domain of attraction (Holling 1973). Environmental disturbances are often vigorous shake-ups, rather than gentle stirrings. Furthermore, there may be no stable equilibrium to return to, but a periodic or chaotic attractor. Consequently, the second conceptual definition is more suitable for ecological systems.
As far as operational definitions in the context of ecosystem models are concerned, a number of indices have been suggested to measure resilience. When the model is explicit, based on differential equations, the most frequently used estimate of resilience is based on the eigenvalues of the linearization of a nonlinear system near an equilibrium point: Pimm and Lawton (1977) measured resilience in terms of the dominant value of the linearization. This is an asymptotic property giving the rate of decay of disturbances as time progresses to infinity. To complement resilience as a measure of the response to disturbances, Neubert and Caswell (1997) proposed indices measuring the extent and duration of the transient growth in models with asymptotically stable equilibria. In the case of individual-based models and cellular automata, resilience is studied with simulations as the time needed after some kind of disturbance to return to its original state (Ortiz and Wolff 2002) or to reach a certain percentage of the previous abundance (Matsinos and Troumbis 2002).
However, the concept of resilience introduced by Holling (1973) stands in contrast to the concept of stability in the strict static sense: defining resilience as how fast the variables return to equilibrium is emphasizing equilibrium conditions, low variability, resistance to and absorption of change, whereas resilience emphasizes the boundary of a stability domain and events far from equilibrium, high variability, and adaptation to change. Resilience is therefore a thoroughly dynamic notion. In this respect, van Coller (1997) showed how techniques from dynamic system theory help us understand the behavior of ecosystem models. The bifurcation diagrams provide a concise way of summarizing the results and can be used to characterize the resilience of models to both parameters and disturbances. Resilience is evaluated as a distance to bifurcation points in Ludwig et al. (1997) and as inversely proportional to the size of attraction domains in van Coller (1997). These measures evaluate the resilience of properties that correspond to attraction domain. They are relevant when the dynamics of the ecosystem model are deterministic. Therefore, the feedback law has to be defined first, and the results only concern this particular feedback law. Furthermore, Bonneuil (2003) shows that the feedback law choice in the well-known predator–prey model may be considered arbitrary because many other laws would have produced a similar behavior. This criticism is all the more pertinent when socio-economic systems are modeled.
We propose an operational definition of resilience that takes into account our ignorance of the laws relating certain controls to the states of the system, and that comes close to Holling’s interpretation. We address the problem from the framework of the viability theory. The concept of resilience depends on: (i) the state of the system, (ii) the objectives (the property to be maintained—in the case of a lake, the resilience of the oligotrophic property or the eutrophic one may be studied; in the case of a farmer population, the resilience of the profitability of their activities may be studied and, in the case of a lake surrounded by a farmer population, the resilience of both oligotrophic property and profitability may be studied), (iii) the anticipated types of disturbances (the stronger the disturbance is, the less resilient any state of the system will be whatever the property under consideration), (iv) the cost associated with the effort that is necessary to restore this property (this cost may be either economic—the amount of money needed to restore the property; or ecological—the length of the period during which the population has to put up with the sight of turbid water in the lake; or both), (v) the control measures that are available, and (vi) the time scale of interest (Carpenter et al. 2001). The challenge is to provide an operational definition that explicitly takes into account these six points. The viability approach (Aubin 1991) deals with dynamic systems under state constraints. This method analyzes the compatibility between the (possibly nondeterministic) dynamics and state constraints. It also determines the set of controls that would prevent the system from violating the state constraints. In this paper, we apply this approach to the definition of resilience. We first describe the evolution of the state of the system (i) influenced by possibly several admissible controls (v) and then governed by possibly nondeterministic dynamics. We next consider the objectives (ii) as state constraints. Then, using the mathematical concept of a viability kernel (Aubin 1991), we highlight the need for anticipation of maintaining a property during the time scale of interest (vi). This analysis also allows us to identify situations where the property may not be maintained. In such cases, we need to refine this analysis to evaluate the cost associated with the effort necessary to maintain or restore this property after a disturbance (iii). So, we extend another mathematical concept, the time of crisis (Doyen and Saint-Pierre 1997) used by Béné et al. (2001) to analyze overexploitation of marine renewable resources. Finally, resilience is evaluated as the inverse of this cost.
With the above in mind, the mathematical formulation of resilience is built in the framework of the viability theory (Aubin 1991).
The model
The dynamics (i) and the controls (v)
The state of the ecological system is described at time t by a vector x(t) in the state space X. This vector may contain several variables: for example, the amount of phosphorus in the lake and the annual phosphorus input. The dynamics depend on the state of the system x(t) and on the different external actions called controls u(t) that can influence the evolution of the system at time t: for instance, banning excessive phosphorus inputs to a lake. A map f associates the variation of x(t) at time t with x(t) and u(t):
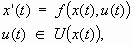 |
[1] |
where u(t) in U(x(t)) is one of the admissible controls when the state of the system is x. The admissible controls are the controls through which an institution (a governmental agency or a private manager) acts on the system. It can choose any of them and it can change from one to another; it also has enough resources to bear the costs associated with these changes.
These dynamics may be nondeterministic if several controls are admissible. If we allow u to be dynamically driven by some feedback law, the dynamics are described by an ordinary differential equation and the evolution of x(t) is deterministic. However, as feedback laws are often arbitrarily chosen (Bonneuil 2003), we do not make further assumptions than u(t) in the set U(x(t)). The solution x(.) depends on the chosen admissible control u(.) which is not known beforehand.
The objectives regarded as state constraints (ii)
The objective is to maintain a system property, for example, to maintain a lake in an oligotrophic state. We consider the map h, which associates with the state of the system, an indicator of this desired property. It holds that, if this indicator belongs to a particular subset, e.g., M, the lake is oligotrophic if the amount of phosphorus in the water is below a given threshold. The set of all states such that this property holds is :
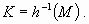 |
[2] |
This set K defines the set of state constraints to be considered in the viability approach.
The objective of maintaining a system property of interest is a rather static assumption that may seem poorly adapted to the changing world we live in: a population that wants an oligotrophic lake at t = 0 may, a few years later, prefer to have an eutrophic lake to use as nutrient sink. However, our aim is not to simulate the evolution of a system over a period of T. The question we attempt to answer is whether a property that is considered a property of interest at t = 0, can be maintained until time T and, if the answer is yes, how can it be maintained and how much will it cost to do so. No doubt the property that will be of interest at a later date will not be exactly the same.
The time scale of interest (vi)
The time scale of interest is defined by T, which may be infinite.
A viability analysis
The viability kernel
The evolution of the system is governed by the dynamics described in Eq. 1. If its state x(0) at t = 0 belongs to K, that means that the property holds at t = 0. The first question that then arises is whether this property can be maintained until T. As several controls may be admissible at each state of the system, there may be many trajectories starting at state x(0) in K satisfying Eq. 1 for t in [0, T]. If one of them remains in K until time T, it means that there exists a control function such that the property holds during the time scale of interest and x(0) is called a viable point. If none of them remains in K until time T, it means that starting at x(0) the system is doomed to lose the property represented by K whatever the controls chosen among the admissible ones and x(0) is a non viable point. The first part of the analysis consists in determining the set gathering all states from which there exists at least one trajectory remaining in K until time T. Aubin (1991) showed the existence of the largest closed viable set in K under the dynamics described by Eq. 1, and called it the "viability kernel" of the set K, denoted Viab(K). This set depends on the dynamics, the set of state constraints, and the set of admissible controls. It is a subset of K that contains, of course, the equilibria but also the states from which there exists one trajectory remaining in K. These trajectories may be governed by permanent external control (viability niche). Regulation through changes in control can also take place. More details are given in Appendix 1.
The cost function (iv), the disturbance (iii), and the resilience
The cost may be socio-economic, ecological, or both. The cost function associates trajectory x(.) with its cost. Usually this cost is calculated by adding the gains and subtracting the expenses at each time step. In our approach, its definition is quite different. The cost is used to evaluate the resilience of one property of the system, so we have to relate its definition to the property under consideration: the cost must measure the capacity of the system to maintain, or at least restore, this property before time T. In other words, the cost of a trajectory must measure a distance between the trajectory followed by the system and the objective imposed on it: maintaining the property or at least restoring it before time T. Such a cost function is used in Béné et al. (2001) in their model dealing with the management of a marine renewable resource, the variable x represents the benefit. The constraint (or the objective) imposed on this variable is to be positive and the cost associated with the trajectory x(.) is the period of negative benefit. As a distance between a trajectory and the objective, the cost function has to satisfy the following two conditions.
First, when the objective is reached , the cost must be null. Therefore, the cost of a trajectory along which the property of interest is maintained is null. Maintaining this property may impose some changes in controls and these changes cost. However, we do not take them into account because these costs are borne by the institution that acts on the system and are thus external; in the same way, Béné et al. (2001) do not take into consideration the cost of the time variation of the fishing effort in the evaluation of the global benefit in the sector.
Second, the cost of a trajectory such that x(T) does not belong to K is infinite. Certainly, the states where the property does not hold are not useless. An eutrophic lake can for example still serve as nutrient sink. However, as far as the evaluation of the resilience of a property is concerned, the objective is to maintain this property or at least restore it before time T. x(T) does not belong to K means that the property has not only been lost but also it has not been restored before the end of the time scale T. Therefore, the cost of such a trajectory must be infinite as the objective is totally missed.
There may be many trajectories starting at x and satisfying Eq. 1 because many controls may be available. The trajectory with minimal cost starting at x is then the best trajectory to follow according to the objective to maintain or restore the property of interest before time T.
Resilience is the capacity of a system to undergo disturbance, so its value also depends on the anticipated disturbance, which is described by a set-value map D that associates state x with set D(x) of all reachable states from x after this disturbance. When a disturbance occurs, the system jumps from a state x to a state y in D(x). The consequence of this jump in terms of the objective to maintain or restore a property of interest, the possible damage, is measured by the possible cost of restoration from the state of the sytem after the jump that is the minimal cost over all trajectories starting at y. The resilience of the system at state x toward a jump to state y is defined as the inverse of this cost. In particular, if the cost starting at y is infinite, the resilience of the system at state x toward a jump to state y is null. This definition of the resilience toward a jump from x to y is summarized by the following equation :
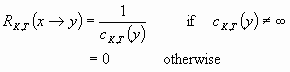 |
[3] |
where cK,T(x) is the minimal cost among all trajectories starting at x and satisfying Eq. 1 and with the convention that the inverse of 0 is infinity.
The anticipated disturbance is defined as a set of attainable states D(x). The precautionary principle leads us to take into account the worst case in the definition of the resilience. So, the resilience of state x facing disturbance D equals the minimal value of the resilience over all jumps from x to y in D(x):
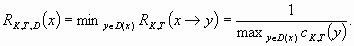 |
[4] |
Lake ecology and economic interpretation
Two states of lakes are usually defined in the literature, a clear-water or oligotrophic state with relatively high economic value of ecosystem services (freshwater, used for irrigation, municipal water supplies, pollution dilution, and recreation) and a turbid-water or eutrophic state. Many lakes have experienced sudden shifts from oligotrophic to eutrophic states. Phosphorus (P) is the most critical nutrient for the eutrophication of lakes (Carpenter et al. 1999 a,b). Excess P is imported to farms in the form of fertilizer and animal feed supplements. P is added to the soil as inorganic fertilizer or manure. Most of the P accumulates in soil, which may then be transported to streams and lakes during runoff events associated with snowmelt or rainstorms. The system under study is composed of the lake and the population that benefits from its services or which provokes excess P inputs as a result of their farming activities. In this simplified model, as far as farmers are concerned, the main issue is ensuring the profitability of their activities. As far as lake managers are concerned, the objective is to maintain the lake in an oligotrophic state. As far as the manager of the whole system is concerned, the crucial issue is, knowing the concentration of phosphorus in the lake and the amount of inputs, can the lake remain oligotrophic and the farming activities remain profitable? After a disturbance, such as an increase of the concentration of phosphorus, is the lake doomed to become eutrophic? Can it become oligotrophic again? If the answer is yes, what are the best actions to undertake and what will be their cost in terms of time spent to recover or in terms of length of the period of negative benefit for farmers?
The operational definition of resilience proposed above provides answers for all these questions.
The model
The dynamics (i) and the controls (v)
To describe the essential dynamics of lake eutrophication we use a simplified version of the model described in Carpenter et al. (1999 a,b), assuming that the sediments are the major source of recycled phosphorus (ignoring the recycling by consumers):
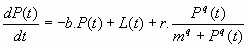 |
[5] |
where P is the amount of phosphorus (mass or concentration) in the water, L is the annual phosphorus input from human activities (mass or concentration per unit time) and b is the proportion of P lost at each time step. The maximum recycling rate of phosphorus (mass or concentration per unit time) is r. The overall recycling rate is assumed to be a sigmoid function of P. The value of P at which recycling reaches half of the maximum rate is m.
Carpenter et al. (1999 a) have derived a bifurcation diagram to associate different types of lakes with sets of parameter values. Fig. 1 shows the bifurcation diagram for the simple lake model in the two-dimensional parameter space (L,P) using parameter values from Janssen and Carpenter (1999): q = 8, r = 1, m = 1 and b takes three different values corresponding to the three types of lakes (b = 0.4 irreversible lake, b = 0.8 hysteretic lake, and b = 2 reversible lake). For b = 2, the low stable equilibrium remains unique as L increases. However, for b = 0.4 and b = 0.8, the stability of the low equilibrium becomes precarious as L approaches L2 because the domain of attraction shrinks; finally, at L = L2, the lower two equilibria disappear and the concentration of phosphorus jumps to the high value. Once the concentration has reached the high equilibrium, it is not easy to reduce it to the lower equilibrium: if L is reduced below L2, the concentration remains at the high equilibrium. For b = 0.8, as L declines below L1, there is a jump down to the low equilibrium that is not reversed as L increases again. This effect is called the “hysteretic effect.” For b = 0.4, jumping down to the lower equilibrium is impossible because L is positive. The jump to the higher equilibrium is irreversible.
As far as the dynamics of the phosphorus inputs are concerned, we assume that the manager can act on their time variation, d L/dt, for example, by enacting constraining laws. For simplicity, we suppose that the derivative of L directly depends on the control u chosen by the manager:
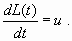 |
[6] |
As modifications of behaviors and changes in agricultural technology and in human and physical infrastructure take time, we consider that the time variation of the phosphorus inputs from human activities is bounded, so u must belong to the closed set of admissible controls:
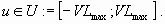 |
[7] |
The objectives regarded as state constraints (ii)
Oligotrophic lakes are characterized by low nutrient inputs, low to moderate levels of plant production, relatively clear water, and a relatively high economic value of ecosystem services. Eutrophic lakes have high nutrient inputs, high plant production, murky water, with problems including anoxia and toxicity, and relatively low value of ecosystem services. So, the lake manager aims to keep it in an oligotrophic state. We assume that an oligotrophic lake becomes eutrophic when the amount of phosphorus in the water increases over some fixed threshold Pmax whatever its type. Consequently, the lake manager’s objective is reached when the positive variable P satisfies:
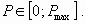 |
[8] |
Certainly, the oligotrophic property of a lake does not only depend on the phosphorus in water, but also on the phosphorus in the mud or in the soil. Ludwig et al. (2003) compared the optimal policies calculated with the one-dimensional model and with the two-dimensional one in which the mud dynamics are taken into account. They underlined the importance of the slow dynamics when the time horizon for optimization is long enough for substantial changes to occur. For simplicity, we neglect the mud and soil dynamics. Nevertheless, the same approach can be applied with two additional variables and a set of state constraints where the oligotrophic property holds, described by conditions imposed not only on the phosphorus in the water but also on the phosphorus in the mud and in the soil.
The farmers’ aim is to ensure the profitability of their activities. We suppose in this simple model that their benefit depends linearly on the inputs of phosphorus. Consequently, their objective is reached when the value of phosphorus inputs from human activities is higher than a given threshold Lmin. We also assume that there is a maximal threshold for phosphorus inputs Lmax fixed by institutions or stakeholders. So, the farmers’ activities are profitable and legal when:
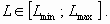 |
[9] |
Equations 5, 6, 7, 8, and 9 can be written synthetically under the formalism of Eq. 1 with x(t): = (L(t),P(t)):
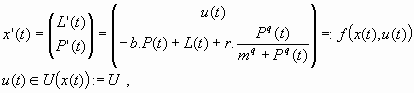 |
[10] |
under the constraints
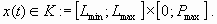 |
[11] |
Like Pmax in the previous paragraph, Lmin may depend on socio-economic variables in a more complex model. Taking it into account would lead to a more complicated shape for the set of states constraints K, but the same approach can be applied because in the viability theory, the only constraint on K is that K has to be closed.
The time scale of interest (vi)
We choose a value of the time scale of interest, T, large enough to see the effects of drastically diminishing the phosphorus inputs. Thus, as the maximal value of allowed phosphorus inputs is Lmax and the maximal decreasing rate is VLmax , the value of T has to satisfy:
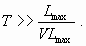 |
[12] |
The cost functions (iv)
We recall that these cost functions are used to evaluate the resilience of the property defined by the set K which, in this application, ensures the profitability of the farmers’ activities and keeps the lake in an oligotrophic state. Consequently, these functions have to satisfy two conditions: first, the cost of a trajectory along which the property is maintained is null; second, the cost of a trajectory such that x(T) does not belong to K is infinite. Furthermore, the trajectory starting at x with minimal cost is the best trajectory to follow according to the objective to maintain or at least restore the property of interest before time T.
In practice, the ways of evaluating the cost of a trajectory x(.) = (L(.),P(.)) satisfying Eq. 10 are numerous and depend on the situation.
If the farmers refuse the slightest negative benefit and if only the time needed by the lake to recover matters, the cost of a trajectory x(.) is infinite if it crosses the line L = Lmin and otherwise is measured by the time spent in an eutrophic state. The time spent by this trajectory outside K is a functional that was introduced by Doyen and Saint-Pierre (1997) and called the “time of crisis.” The function γ that associates x with the minimal cost over all trajectories starting at x is then defined by:
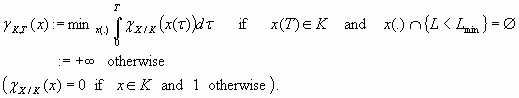 |
[13] |
In the second cost function, the farmers’ objectives do not totally prevail over the ecological ones. The cost function is made up of two terms: the first term, which corresponds to the ecological cost, is the same as in the cost function γ; the second one, which is an economic cost, measures the time duration of the period of negative profits weighted by the norm of these negative profits. The first term is multiplied by a factor c
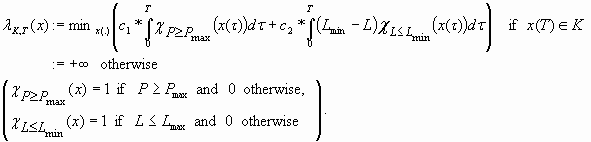 |
[14] |
Actually, the first cost function equals the second one with c
The disturbance (iii)
In this simple example, we consider disturbances Dα corresponding to a sudden increase of the concentration of phosphorus in the lake. α represents the maximal intensity of the anticipated disturbance. Such a disturbance modifies the concentration of phosphorus P but leaves the amount of inputs L unchanged. If a disturbance Dα occurs when the state of the system is x, it will jump to a state y belonging to set Dα(x):
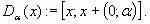 |
[15] |
In the simplest case (when the cost functional is the time of crisis γ), the viability kernel, the cost function, and the resilience level curves can be solved analytically, and the plots are obtained by numerical integration of differential equations. As far as the second cost functional λ is concerned, we use the viability algorithm (Saint-Pierre 1994) to compute the resilience values. Viability Kernels
The first question that arises is whether the evolution (Eq. 10) is compatible with the objective represented by the set K (Eq. 11): ensuring the profitability of the farmers’ activities and keeping the lake in an oligotrophic state. In other words, our aim is to determine the levels of phosphorus inputs and phosphorus in the water, (L,P), belonging to the set K such that there exists a trajectory starting from them and remaining in K until time T. These trajectories are called viable trajectories because the objective is completely met when such trajectories are followed. We first identify viable stationary points, then viability niches, and finally we compute the whole viability kernel that gathers all states from which there exists at least one trajectory satisfying Eq. 10 remaining in K until time T. The viability kernel is then smaller than the set of state constraints K.
Viable stationary points
The stationary points correspond to d L/dt = 0 and d P/dt = 0. These are viable when they belong to K. Thus, the viability kernel of the set of state constraints K contains the equilibria xeq = (Leq,Peq) such that
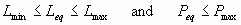 |
[16] |
The set of viable equilibria corresponds to the line EQ in Fig. 2. The solid part of the line contains the stable equilibria, the dashed part the unstable ones.
Viability niches
The viability niches correspond to initial phosphorus inputs and phosphorus in water such that the resulting evolution with a permanent external control u(t) = u0 remains in K. Clearly viable equilibria are part of viability niches. As d L/dt = u0 and L must be bounded for a trajectory to be viable, the only non empty viability niche is the viability niche of u0 = 0, N(0). u(t) = 0 implies L(t) = L(0) = L0 is constant and the variation of P(t) from P(0) to one of the possible three steady states Peq(L0) is monotonous. Thus, if Peq(L0) belongs to K, the intersection between the attraction domain of Peq(L0) and K belongs to N(0). N(0) is the union for L0 = Lmin to Lmax of such intersections. This viability niche is colored light gray in Fig. 2.
The viability kernel
The viability kernel differs from the niches in that, for the kernel, regulation through changes in control can occur, thus allowing the viability to be increased. Such changes are limited to the set of admissible controls: the intensity of the time variation of the phosphorus inputs must be smaller than VLmax, modeling the resistance to change and the rigidity of the decision. When VLmax tends toward 0, the viability kernel tends toward the viability niche N(0) in accordance with Carpenter et al. (1999a). An increase of VLmax allows a greater decrease in phosphorus inputs from human activities, so the surface of the viability kernel increases with VLmax (Fig. 3).
We can distinguish three qualitative configurations for the viability kernel depending on the parameters (Lmin,Lmax,Pmax). These three cases are illustrated in Fig. 4 for an irreversible lake, in Fig. 5 for an hysteretic lake, and in Fig. 6 for a reversible lake.
Case 1 is global viability. This is the most favorable case because the viability kernel equals the whole set of state constraints, Viab(K) = K. This situation occurs when Pmax is larger than all the P-values of the equilibria associated with Lmax. This statement means that, whenever the maximal phosphorus input is low enough or, in a symmetric way, whenever the threshold of eutrophication is high enough compared with the P-values of the equilibria associated with Lmax, viability holds everywhere. This global viability is represented in Fig. 6 in the case of a reversible lake.
Case 2 is partial viability. The viability kernel is a strict but non-empty subset of K. This case occurs when Pmax is strictly smaller than the highest P-values of the equilibria associated with Lmax and bigger than the smallest P-values of the equilibria associated with Lmin. The curve g defines the right boundary of the viability kernel and corresponds to a trajectory satisfying d L/dt = -VLmax (Fig. 4, Fig. 5, and Fig. 6). This curve represents the states of the system where it is essential to change the control and choose u = -VLmax, the maximal decreasing rate of phosphorus inputs. The set K/Viab(K) contains situations where the values of the phosphorus inputs and the phosphorus in water are too high to maintain the trajectory in K until time T regardless of the control choices.
In case 3, the viability kernel is empty. This situation occurs when Pmax is strictly smaller than the smallest P-values of the equilibria associated with Lmin. In particular, no equilibrium belongs to K because an equilibrium located in K always belongs to Viab(K). This situation most likely occurs when the state of the lake is irreversible (Fig. 4 ).
Numerical results of the next two subsections are computed with fixed values of the following parameters: Lmin = 0.1, Lmax = 1, Pmax = 0.5, VLmax = 0.1 and T = 50.
Cost FunctionsWe noted in the preceding section the possible existence of partial viability. From the states belonging to K/Viab(K), the dynamics cause the system to leave the domain of constraints regardless of the controls: the lake is doomed to become eutrophic, or the benefit is doomed to become negative. The property of interest cannot be maintained. The question that then arises is whether a trajectory starting from x in K/Viab(K) exists, such that this property is restored before time T. If the answer is yes, the cost of following this trajectory is evaluated by cost functions that represent measures of the distance between it and an ideal trajectory along which the property would have been maintained (this ideal trajectory is thus contained in K). The minimal cost over all trajectories starting at state x of the system and satisfying Eq. 10 is the most optimistic prediction of the future cost when the present state of the system is x. This cost depends on the viewpoint of the manager. We choose to study the values of two different arbitrary cost functions defined by Eq. 13 and Eq. 14.
Trajectories from K/Viab(K)
Figure 7 shows trajectories from different states that are in K but not in Viab(K) and governed by controls such that:
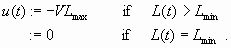 |
[17] |
All trajectories leave K sooner or later because they start from states in K but outside Viab(K). However, some trajectories reach Viab(K) before time T (Fig. 7, plain lines) and thus their cost can be finite. Other trajectories converge toward an equilibrium outside K (Fig. 7a dotted lines), their costs are infinite whatever the cost function γ or λ because the property of interest cannot be restored.
Level curves of the cost functions
Common features
The two cost functions divide the space X into three areas:
(i) the set of states for which the cost is zero (gray area in Figs. 8–9). This set equals Viab(K) for all cost functions;
(ii) the set of states for which the cost is finite. In this area, the level curves do not merge with each other; and
(iii) the set of states for which the cost is infinite (for instance, the states for which the optimal trajectories are drawn as dotted lines in Fig. 7a). This set contains the states from which the property cannot be restored regardless of the cost function. It is shaded gray in Fig. 8 and white in Fig. 9.
The cost function γ
In the case of γ, the time of crisis reaches its minimum for the trajectories controlled by Eq. 17 shown in Fig. 7 (mathematical proof in Appendix 2): the optimal policy is to decrease the inputs down to Lmin with the maximal admissible velocity (-VLmax) and then to keep L = Lmin constant because the farmers cannot support the slightest negative benefit. It is worth noting that, depending on the starting state, this policy may succeed in restoring the oligotrophic property (see trajectories drawn as solid lines in Fig. 7) or may fail (see trajectories drawn as dotted lines in Fig. 7a). The time of crisis can be evaluated by numerical integration along such trajectories. The plots of the level curves of this time of crisis (i.e., the lines along which the time of crisis is constant) are drawn in Fig. 8. The optimal trajectories and the time of crisis level curves merge inside K/Viab(K) because the time of crisis measures the time spent outside K.
The cost function λ
For the second cost function, λ, we cannot find the optimal trajectories analytically. We use the characterization of the cost function map in terms of the boundary of a viability kernel described by Doyen and Saint-Pierre (1997) and then compute the values of the cost function λ thanks to the viability algorithm (Saint-Pierre 1994). We evaluate the values of this cost function for two sets of parameter values: c1 = 0.3, c2 = 17, and c1 = 1.5, c2 = 5. In the second case, the importance of the ecological issue is larger compared with the economic one than in the first case. The level curves of the cost function λ are plotted in Fig. 9 and Fig.10 displays the 3-D graph of this cost function. Contrary to the cost function γ, the area L<Lmin is not forbidden. The optimal policy is to decrease the inputs down to L0<Lmin for the oligotrophic property to be restored faster and then to increase the inputs to Lmin for the farmers’ benefit to be positive again. The value of L0, which depends on the starting state of the system, is the answer for the tradeoff between the economic and ecological issues. The more important the ecological issue is compared with the economic one, the smaller L0 is.
Disturbance Cost and Resilience
The cost functions associate a state of the system with the minimal cost necessary to restore the property of interest. When a disturbance occurs, the system jumps from state x to state y. The minimal cost associated with y provides a measure of the possible damage caused by the jump in terms of the capacity to maintain, or at least restore, this property. The disturbances we consider are a sudden increase of the concentration of phosphorus in the water. Following the precautionary principle, their cost is the maximal cost engendered by the system jump from state x to state y in Dα (Eq. 15). The inverse of this cost provides the measure of resilience we propose.
Disturbance cost
In this simple example, whatever the state of the system x and the cost function (γ or λ), the maximal cost is caused by the jump of maximal magnitude. Consequently,
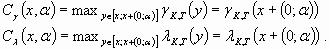 |
[18] |
Level curves of the resilience
Main features
Resilience is defined as the inverse of the disturbance cost and depends on the intensity of the disturbance, α and on the cost functional choice, γ or λ. From Eq. 18, the resilience values are obtained by:
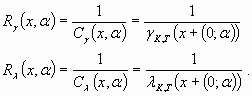 |
[19] |
We evaluate the resilience values for each cost function and for one value of the maximal magnitude of the anticipated disturbance. The level curves of the resilience are plotted in Fig. 11 for an irreversible lake and in Fig. 12 for an hysteretic lake.
The resilience functions divide the space into three areas:
(i) the areas shaded light gray correspond to infinite resilience: the state of the system remains in Viab(K) even if a disturbance of the anticipated magnitude occurs,
(ii) resilience is finite between the level curves: following a disturbance of anticipated magnitude, the lake may become eutrophic instantaneously or later. However, the oligotrophic state can be restored before time T,
(iii) resilience is null for the states belonging to the dark gray areas: these areas contain the states from which a disturbance of the anticipated magnitude may make the system jump to a state where it is doomed to leave K and cannot return before time T.
Comments
According to our operational definition, the resilience value depends on:
(i) the state of the system: Fig. 11 and Fig. 12 show the level curves of the resilience that is the resilience value according to the state of the system,
(ii) the objectives represented by the set K,
(iii) the intensity of the anticipated disturbance: Fig. 13 plots the resilience value of the state with coordinate L = 0.2 and P = 0.25 according to the magnitude of the disturbance α. For low values of α, resilience is infinite. Thus, as α increases, resilience decreases,
(iv) the way of evaluating the costs: the comparison between the plots in Fig. 11 or in Fig. 12 highlights the influence of the choice of the cost function. Two states may belong to the same level curve of the resilience for one cost function choice and to different level curves for another choice. In the first case, these states have identical resilience values, in the second case, one state is more resilient than the other,
(v) the admissible controls represented by the set {U(x)}x, and
(vi) the time scale of interest, T, which do not vary in this subsection but strongly influence the shape of the viability kernel (see the subsection entitled “Viability kernels”).
A large part of ecological theory focuses on asymptotic behavior (Hastings et al. 1993, Ludwig et al. 1993) and the static viewpoint inspires many operational definitions of resilience: Neubert and Caswell (1997) provide two indices for measuring the extent and duration of transient growth caused by a perturbation from an asymptotically stable equilibrium. In the case of individual-based models and cellular automata, resilience is studied with simulations as the time needed after some kind of disturbance to reach a certain percentage of the previous abundance (Matsinos and Troumbis 2002 ; Ortiz and Wolff 2002).
However, nonlinear dynamics do conventionally characterize ecosystem models and nonlinearity is notorious for producing long and/or large transients. Resilience emphasizes events far from equilibrium. In this respect, van Coller (1997) and Ludwig et al. (1997) put the emphasis on the boundary of a stability domain: they evaluate resilience in terms of distance to bifurcation points or size of attraction domains. Their approach applies consistently to dynamic models described by differential equations. In this paper, we aim at going further and taking into account our ignorance of the laws relating certain controls to the states of the system.
We have proposed a new mathematical formulation of resilience in the framework of the viability theory. The main purpose of this new approach is to provide an operational definition of resilience that takes into account the variety of dynamics available to the system and that comes close to Holling’s interpretation. First, we have described the dynamics of the system, allowing several time variations depending on the chosen control at each time step. Second, we have studied the resilience of one property of interest: we have identified the states where this property holds as a set of state constraints in which the trajectory has to be contained for the property to be maintained during the time scale of interest. Third, in contrast to the static viewpoint, we have focused on the notion of trajectory. More precisely, we have focused on the set of trajectories starting from any state of the system and associated with different control functions. At each trajectory, we have associated a cost that is either null if the property is maintained along the trajectory or that represents the cost of restoration if the property is lost and then restored. This cost is infinite if the property cannot be restored. Fourth, we have defined the resilience value at one state of the system as the inverse of the minimal cost over all trajectories starting at the state to which the system has jumped following the disturbance.
To apply our definition of resilience, we have revisited a highly simplified classical model of lake eutrophication. This case study demonstrates that our definition, together with the viability algorithm, is operational. Furthermore, it shows that our definition takes into account all the parameters used in conceptual definitions of resilience: the state of the system, the objectives (the property to be maintained), the anticipated types of disturbances, the cost associated with the effort necessary for restoring this property, the control measures that are available, and the time scale of interest (Carpenter et al. 2001). Notwithstanding that this simplified model falls short of accurately representing the complex eutrophication process, it is nonetheless an adequate proxy for straightforward testing of the validity of our new definition of resilience.
This work suggests a number of interesting open problems. For example, theoretical and experimental ecologists have studied how resilience is affected by ecosystem characteristics, including energy flow (DeAngelis 1980), nutrient loads and nutrient cycling (Steinman et al. 1991), food chain length (Cottingham and Carpenter 1994), and food web connectivity (Armstrong 1982). It would be interesting to see how these properties affect our definition of resilience.
Our results concern the mathematical definition of resilience and its calculation from mathematical models. The comparison between the calculated trajectories and those obtained by historical empirical data would constitute another relevant way of validating the model. Thus, mathematical concepts and tools provided by viability theory can be helpful in constructing better models. Moreover, our results have implications for empirical studies: our approach provides the controls to be used to follow the trajectory with the minimal cost. Applying these controls on real ecosystems may improve their recovery.
However, it is clear that the model adopted in this study is stylized and built on simplistic assumptions. Some of them have been made for the sake of simplicity and are easy to relax as the viability theory by itself imposes lesser constraints. We have assumed that the oligotrophic property of a lake only depends on the phosphorus in water, and we have also assumed that Lmin yields zero profits and any L larger than Lmin would yield positive profits. Consequently, the set of state constraints, K, is a rectangle. We can relax these assumptions by taking into account the slow dynamics of the phosphorus in the mud and in the soil, and the socio-economic variables upon which the farmers’ profits may very well depend. This more complex model would lead to a less regular shape for the set of states constraints K, which would be described by conditions imposed not only on the inputs of phosphorus and the phosphorus in the water but also on the other variables. Nevertheless, the same approach can be applied to this more generic framework because, in the viability theory, the only constraint on K is that K must be closed.
In the mathematical formulation of the definition of resilience, this set of state constraints K does not depend on t. Nevertheless, the property considered as a property of interest at t = 0 may depend on t, for example, having an increasing benefit. The trajectory must then remain in K(t) for the property to be maintained. In such simple cases as the increasing benefit, it is easy to modify the dynamics in such a way that K(t) becomes constant. However, in more complex cases, when the dependence of K(t) on t cannot be removed, the viability analysis is beyond the scope of this article.
Our cost functions also suffer from oversimplifications. Only simple cost functions can be studied analytically. However, the characterization of the minimal cost values in terms of the boundary of a viability kernel allows us to compute the minimal cost of more complex functions. The assumptions imposed on functions in the viability theory are described in Appendix 1. Usual cost functions satisfy them.
It is also worth noting that real ecosystems are seldom if ever subject to simple, temporally isolated disturbances. Our analysis thus deliberately ignores the effect of continual stochastic disturbances, but relies on the hope that the deterministic results will shed light on the stochastic case. Developing a definition of resilience in the framework of viability theory for stochastic systems would enhance the accuracy of using these mathematical concepts to evaluate resilience in ecosystem models. We may address this issue in the framework of a dynamic game (Aubin 1997). The dynamics of the system at state x would not only be influenced by controls u in U(x), which can be chosen, but also by disturbances v in V(x) imposed by nature in an independent way for an unknown purpose. The anticipated features of the disturbances would be described by the conditions imposed on the set V(x) and starting at a state x, the trajectory would depend on the control function u(.) and on the disturbance function v(.). The relevant cost at state x would be similar to the one presented in this paper: the maximum overall anticipated disturbance functions v(.) of the minimum overall available control functions u(.) of the cost associated with the trajectory governed by u(.) and v(.).
Finally, we have applied our definition to a classical simple model of lake eutrophication because our intent was to present our new method and we thought that this would be easier within a well known context. Thus, we have described P-dynamics by the sigmoid function and we have used parameter values taken from literature. However, there are great difficulties in obtaining accurate estimates for the parameters involved in the P and L dynamics (Ludwig et al. 2003) and, consequently, the trajectories on which we have based our definition may be biased. Instead of giving fixed values to each parameter, it would be more accurate to assume that their value at each time step lies within two fixed bounds. This way, the system response to an external action (a control) would not be deterministic. Such a model could be developed in the framework of the viability theory because this theory allows at each time step multiple velocities. There would be many trajectories associated with one control function, depending on the parameter values, and our ignorance of the parameter values could be treated as the disturbance functions of the previous paragraph. Actually, the viability approach does not impose a perfect knowledge of the system dynamics, the velocities may belong to sets and the only constraint imposed on the sets of velocities is that they must be convex.
We have pointed out the advantages and the actual limits of the viability approach. We suggest that this approach is a fruitful framework to define resilience and more generally to address some of the issues encountered in ecosystem management.
Responses to this article are invited. If accepted for publication, your response will be hyperlinked to the article. To submit a comment, follow this link. To read comments already accepted, follow this link.
We thank Isabelle Alvarez , Frédéric Amblard, Noël Bonneuil, Guillaume Deffuant, François Goreaud, and Jean Martin for their constructive comments and very helpful suggestions on an earlier version of this manuscript. We thank Patrick Saint-Pierre for the results of the viability algorithm computed by his software.
Armstrong, R. A. 1982. The effects of connectivity on community stability. American naturalist 120:391–402.
Arrow, K., B. Bolin, R. Costanza, P. Dasgupta, C. Folke, C. S. Holling, B.-O. Janssen, S. Levin, K.-G. Mäler, C. Perrings, and D. Pimentel. 1995. Economic growth, carrying capacity, and the environment. Science 268:250–251.
Aubin, J. P. 1991. Viability theory. Birkhäuser, Boston, Massachussetts, USA.
Aubin, J. P. 1997. Dynamic economic theory. Springer-Verlag, Berlin, Germany.
Béné, C., L. Doyen, and D. Gabay. 2001. A viability analysis for a bio-economic model. Ecological Economics 36:385–396.
Bonneuil N. 2003. Making ecosystem models viable. Bulletin of Mathematical Biology 65(6):1081–1094.
Carpenter, S. R., W. A. Brock, and P. C. Hanson. 1999b. Ecological and social dynamics in simple models of ecosystem management. Conservation Ecology 3:4. (Online.) URL: http://www.consecol.org/vol3/iss2/art4/index.html.
Carpenter, S. R., D. Ludwig, and W. A. Brock. 1999a. Management of eutrophication for lakes subject to potentially irreversible change. Ecological Applications 9:751–771.
Carpenter, S. R., B. Walker, J. Anderies, and N. Abel. 2001. From metaphor to measurement: resilience of what to what? Ecosystems 4:765–781.
Cottingham, K. L., and S. R. Carpenter. 1994. Predictive indices of ecosystem resilience in models of north temperate lakes. Ecology 75:2127–2138.
DeAngelis, D. L. 1980. Energy flow, nutrient cycling, and ecosystem resilience. Ecology 61:764–771.
Doyen, L., and P. Saint-Pierre 1997. Scale of viability and minimal time of crisis. Journal of Set-Valued Analysis 5:227–246.
Gunderson, L., and C. S. Holling. 2002. Panarchy : understanding transformations in human and natural systems. Island Press, Washington, D.C., USA.
Hastings, A., C. L. Hom, S. Ellner, P. Turchin, and H. C. J. Godfray. 1993. Chaos in ecology: is Mother Nature a strange attractor? Annual Reviews of Ecology and Systematics 24:1–33.
Holling, C. S. 1973. Resilience and stability of ecological systems. Annual Reviews Ecology and Systematics 4:1–23.
Janssen, M., and S. R. Carpenter. 1999. Managing the resilience of lakes: a multi-agent modeling approach. Conservation Ecology 3:15. (Online.) URL: http://www.consecol.org/vol3/iss2/art15/index.html.
Ludwig, D., S. Carpenter, and W. Brock. 2003. Optimal phosphorus loading for a potentially eutrophic lake. Ecological Applications 13:1135–1152.
Ludwig, D., R. Hilborn, and C. Walters. 1993. Uncertainty, resource exploitation, and conservation : lessons from history. Science 260:17–36.
Ludwig, D., B. Walker, and C. S. Holling. 1997. Sustainability, stability and resilience. Conservation Ecology 1:7. (Online.) URL: http://www.consecol.org/vol1/iss1/art7/index.html.
Matsinos, Y. G., and A. Y. Troumbis. 2002. Modeling competition, dispersal and effects of disturbance in the dynamics of a grassland community using a cellular automaton. Ecological Modelling 149:71–83.
Neubert, M. G., and H. Caswell. 1997. Alternatives to resilience for measuring the responses of ecological systems to perturbations. Ecology 78:653–665.
Odum, E. P. 1993. Ecology and our endangered life support system. Sinauer Associates, Sunderland, Massachusetts, USA.
Ortiz, M., and M. Wolff. 2002. Dynamical simulation of mass-balance trophic models for benthic communities of north-central Chile: assessment of resilience time under alternative management scenarios. Ecological Modelling 148:277–291.
Pimm, S. L. 1984. The complexity and stability of ecosystems. Nature 327:321–326.
Pimm, S. L. 1991. The balance of nature? University of Chicago Press, Chicago, Illinois, USA.
Pimm, S. L., and J. H. Lawton. 1977. Number of trophic levels in ecological communities. Nature 268:329–331.
Saint-Pierre, P. 1994. Approximation of the viability kernel. Applied Mathematics & Optimisation 29:187–209.
Steinman, A. D., P. J. Mulholland, A. V. Palumbo, T. F. Flum, and D. L. DeAngelis. 1991. Resilience of lotic ecosystems to a light-elimination disturbance. Ecology 72:1299–1313.
van Coller, L. 1997. Automated techniques for the qualitative analysis of ecological models : continuous models. Conservation Ecology 1:5. (Online.) URL: http://www.consecol.org/vol1/iss1/art5/index.html.
Address of Correspondent:
Sophie Martin
Laboratoire d'Ingénierie pour les Systčmes Complexes,
Cemagref, Groupement de Clermont-Ferrand,
24 avenue des Landais,
BP50085 F-63172 Aubičre Cedex, France
Phone: 33.4.73.44.06.87
Fax: 33.4.73.44.06.97
sophie.martin@cemagref.fr
![]()