|
|
|
Copyright © 2004 by the author(s). Published here under license by The Resilience Alliance.
Go to the pdf version of this article.
The following is the established format for referencing this article:
Walker, B. and J. A. Meyers. 2004. Thresholds in ecological and social–ecological systems: a developing database. Ecology and Society 9(2): 3. [online] URL: http://www.ecologyandsociety.org/vol9/iss2/art3/
A version of this article in which text, figures, tables, and appendices are separate files may be found by following this link.
Synthesis Thresholds in Ecological and Social–Ecological Systems: a Developing Database Brian Walker1 and Jacqueline A. Meyers
1CSIRO Sustainable Ecosystems
- Abstract
- Introduction
- The Developing Database
- Examples: Categories of Thresholds
- Class 1. No linkage, externally driven change in ecological or social systems
- Class 2. No linkage, internally driven change in the ecological or social systems
- Class 3: Linked social–ecological systems, with a threshold change in only one system
- Class 4: Linked social–ecological systems with reciprocal influences, but a shift in only one system
- Class 5: Linked social–ecological systems with reciprocal influences, shifts in both the ecological and social systems
- Some Initial Observations on Thresholds
- Conclusions
- Responses to this Article
- Acknowledgments
- Literature Cited
Increasing interest in regime shifts in ecological and linked social–ecological systems (SESs) has placed a strong focus on the thresholds of change. However, research into this topic has been hampered by a lack of empirical data. This paper describes a developing database established to address this need. The database is freely available and comprises a set of summarized published examples and a searchable bibliographic database of publications on the topic. Thresholds in the database are characterized in terms of a standardized set of 24 descriptors, including the variables along which they occur, the variables that change, and the factors that have driven the change. Readers are encouraged to contribute new examples. Examples range from conceptual models to empirical evidence. The former predominate in the literature and, although they make valuable contributions and will continue to be included, the intention is build up the number of examples based on data. Examples are presented in terms of whether the threshold occurs in the ecological system, the social system, or both, and the direction of interactions between systems. The paper concludes with some initial observations on thresholds based on the examples included so far, and poses some questions for future research. Research on a typology of thresholds is a priority topic in the emerging area of “sustainability science” and it requires a rich database of empirical data.
KEY WORDS: alternate states, database, ecosystems, regime shifts, resilience, social–ecological systems, thresholds.
Published: July 22, 2004
C. S. Holling’s (1973) seminal paper on resilience sparked the beginning of a growing interest in the ability of ecosystems to absorb and respond to disturbance. Shortly after, May (1977) published another well-known paper on thresholds and breakpoints in ecosystems with a multiplicity of stable states. Many others have followed and recent reviews (Beisner et al. 2003, Scheffer and Carpenter 2003, Scheffer et al. 2001, Muradian 2001) are evidence that it remains an active and advancing area of research. The work has focused attention on the notion of multiple states, placing an emphasis on the thresholds between them. Attempts to analyze the nature of such thresholds have been hampered by a lack of readily available empirical data. It was recognition of this lack of data, at a meeting of the “Robustness” program of the Santa Fe Institute (http://discuss.santafe.edu/robustness), that led to the development of the database described in this paper.
The term “alternate states” is commonly used to describe the phenomenon whereby systems can exhibit a big change from one kind of regime to another. However, it is important at the outset to distinguish between examples of what truly are alternate stable states—i.e., two or more stable point attractors separated by unstable thresholds—and the various other kinds of big changes that systems may exhibit (cf. Carpenter 2003). To avoid any confusion over terminology, we prefer to follow Scheffer and Carpenter’s (2003) use of the terms “regime shift” and “multiple regimes” and include in the database all the various kinds of thresholds and multiple system regimes that occur, including alternate stable states.
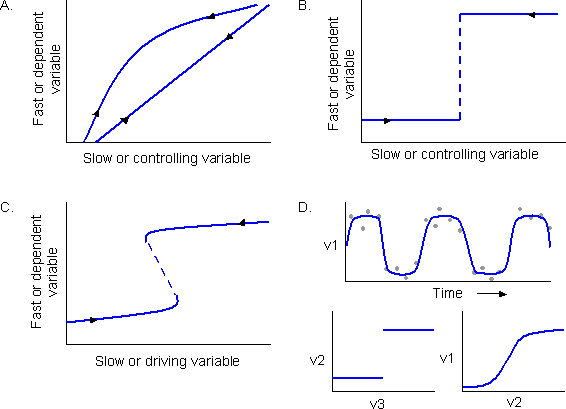
Distinguishing between systems with alternate attractors and those that, although they might sometimes exist in different regimes, have only a single attractor, is a primary reason for this database. The distinction is important because the management and governance implications can be profound. All kinds of regime shifts are included in the database in order that we may learn how to identify and distinguish between them. Four of the main kinds are depicted in Fig. 1.
|
Fig. 1. Possible kinds of changes in systems: (A) no regime shift, changes are continuous and smoothly reversible; (B) regime shift, but there are no alternate attractors (for example, the changes between ice and water in response to temperature); (C) regime shift with alternate attractors; (D) a class of regime shift involving three variables (for example, v1 = salmon stocks, v2 = temperature, and v3 = ocean currents), with no feedback from the dependent (v1) to the slow variable (v3).
|
A regime shift involving alternate stable states occurs when a threshold level of a controlling variable in a system is passed, such that the nature and extent of feedbacks change, resulting in a change of direction (the trajectory) of the system itself. A shift occurs when internal processes of the system (rates of birth, mortality, growth, consumption, decomposition, leaching, etc.) have changed and the state of the system (defined by the amounts of the state variables) begins to change in a different direction, toward a different attractor. In some cases, crossing the threshold brings about a sudden, large, and dramatic change in the responding state variables; for example, the shift from clear to turbid water in lake systems (Meijer 2000, Carpenter 2003). In others, such as a change from a grassy to a shrub-dominated rangeland (Ludwig et al. 1997), the response in the state variables is more gradual but, nevertheless, once the threshold has been passed, the feedbacks have changed and the dynamics of the system shift from one basin of attraction to another. At the shrub density threshold, the feedback from shrubs to grass changes to the extent that there is insufficient grass to enable fire to spread and burn the shrub layer.
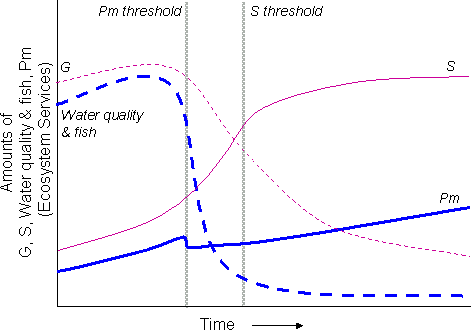
The term “flip” is sometimes used to describe such a shift. However, this term is misleading because it can be used to imply either a rapid, large change in system structure (as in the lakes example), or a sudden change in system feedbacks. It needs to be made clear that, even though crossing a threshold results in a sudden change in system feedbacks, the consequent changes in state variables can range from being fast (as in lakes) to slow (as in rangelands), as illustrated in Fig. 2. Hence we prefer the term regime shift.
|
Fig. 2. Stylized trajectories through time of the fast (---) and slow (____) variables in lakes (thick blue lines) and rangelands (thin red lines) under high levels of phosphate inflow (lakes) and grazing (rangelands). In lakes, there is a very rapid change (“flip”) in water quality as the threshold is passed and an associated rapid decline in water quality and fish. The amount of phosphate in the mud (Pm) shows a short-term decline as phosphate becomes soluble and is lost from the mud to the water above. In rangelands, although the feedbacks change instantly at the threshold (as in Fig. 1c), the lag effect in shrub growth rate results in slow changes of the variables (G = grass; S = shrubs).
|
In the same way that an ecological or physical-environmental system can undergo a regime shift, so too can a social system. Changes may result in different patterns or kinds of social behavior and these functional changes may lead to or be associated with shifts in social structure—such as population numbers. In linked social–ecological systems (SESs), changes in one system may feed back as drivers to alter variables in the other, causing a regime shift in that system. The effect may be one-way, resulting in a regime shift in only one system, or there may be a two-way interaction, resulting in regime shifts in both the ecosystem and society.
Given the array of possible regime shifts, we adopt a broad definition of a threshold as a breakpoint between two regimes of a system. It is intended to be inclusive, to provide information and context for analysis of different kinds of thresholds and different kinds of regime shifts.
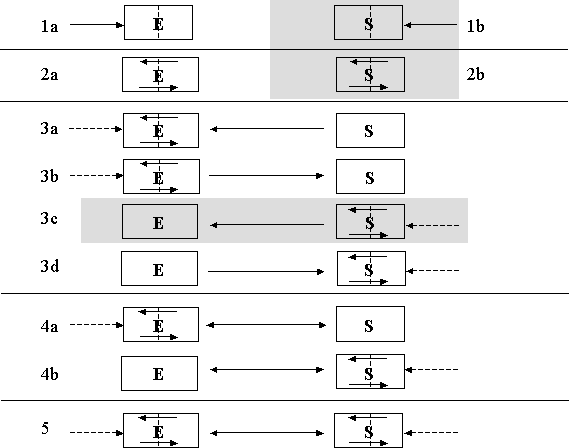
The database focuses on ecological and linked SESs, in particular those that exhibit thresholds in relation to the use of ecosystems in natural resource management. Purely social system thresholds, that are unconnected to the dynamics of a natural system, are not included in the database. Nor are social–ecological examples that describe a change in the social system, but do not identify a change in the ecosystem. Figure 3 shows all the possible combinations of ecological and social interactions and highlights the categories not included in the database. The database does not contain raw data, but it does provide a summary of the threshold-related aspects of the publications and it points to the original data sources.
|
Fig. 3. All the possible interactions between social (S) and ecological (E) systems in relation to threshold shifts. Systems that have undergone a threshold shift to an alternate regime are split with a dashed line. The arrows within the boxes indicate that feedback mechanisms operate within the system. The arrows connecting the social and ecological systems show the direction of interaction between the systems in the development of regime shifts. Dashed arrows indicate that external influences may or may not contribute to the regime shift. The shaded categories are not included in the database, but are shown here for completion.
|
The database was established as a joint activity of the Santa Fe Institute and the Resilience Alliance (a multidisciplinary consortium of research groups from around the world with interests in the dynamics of linked SESs). Users are encouraged to contribute to the database, as its value and usefulness will increase as the number and diversity of examples grows. It is freely available and is accessible under “Thresholds Database” on the Resilience Alliance website (http://www.resalliance.org). It consists of two components: a set of summarized published examples of thresholds and regime shifts in ecological and SESs and a searchable bibliographic database of publications on the topic.
In characterizing the examples, emphasis is placed on describing the threshold: the variables along which the threshold occurs, the variables that change as a consequence of the shift, and the factors that have driven the change. Where relevant, governance or management practices involved in the change are also described. The categories used to describe each example are listed in Table 1. Where appropriate, drop-down menus simplify and standardize the information. Search and sort functions make it simple to find examples on topics of interest. Information in the database can be downloaded into an Excel® file.
|
Table 1. The categories used in the Thresholds Database to describe each example.
The examples in the database range from conceptual models to empirical evidence, but the primary aim is to collect as many examples with empirical data as possible. This will provide researchers with a tool to aid analyses of thresholds. Few studies collected so far have tested a prior hypothesis about a threshold (examples include those by Scheffer et al. 2003, Pech et al. 1992, Robblee et al. 1991). More often, regime shifts have been observed and reported after the shift has occurred. In some cases, retrospective studies have been conducted where measurements taken before the shift were by chance available and could be compared with data collected after the shift: for example, Higgins et al. (2002) and Hare and Mantua (2000) who, respectively, collected 31 physical time series of atmospheric and oceanic processes and 69 biological time series of ocean fauna over 33 years. The data showed that climate oscillations resulted in a shift in water temperature across the North Pacific Ocean, with varying responses in fauna. Many other examples are based on descriptive data, where a shift in state has been observed, but not measured. In other examples, there is a strong suggestion of a shift having occurred but the threshold point at which the system feedback changes direction is not apparent. In still other examples, data from areas that have already undergone a shift have been compared with data from similar environments that have not, as in the study by Sarre et al. (1995) where population persistence of lizards from vegetation remnants was compared with that of lizards from sites in continuous woodland.
To aid description and facilitate database searches aimed at exploring particular questions about thresholds and the nature of the systems in which they occur, the examples have been organized into five classes, according to whether the regime shift has occurred in the ecological, social or linked SES. These five classes are further divided into eleven categories based on the direction and impact of the interactions between the systems (Fig. 3). Thresholds in classes 1 and 2 are in either an ecological or a social system, with no linkage between systems influencing the thresholds. The driving force of change comes either from outside (Fig. 3, 1a, 1b) or from within the system (Fig. 3, 2a, 2b). Thresholds in classes 3–5 are all in some way affected by links between the ecological and social systems. In class 3, one system influences the other, but there is a threshold change in only one system and no feedback to the other. In class 4, there is reciprocal influence, but with a regime shift in only one system. Finally, in class 5, there is a reciprocal influence and a regime shift in both systems. As mentioned previously, the database does not include social examples in classes 1 and 2, or the social shift that brings about change (but not a regime shift) in the ecological system (3c). Some illustrative examples of each category in the database follow. Class 1. No linkage, externally driven change in ecological or social systemsCategory 1a. Shift in the ecological system, driven by an environmental event from outside, with no impact from societyMajor environmental events such as asteroid impacts, volcanic eruptions, earthquakes, or extreme weather events fit into this category, but are only included if there is a threshold effect associated with the event. At first, the drivers (i.e., the events) may appear to be obvious but Zinsmeister (1998) suggests the true drivers may be more subtle and the pathways to the regime shift more complex. Rather, he suggests, the Earth’s biosphere tends to be resilient to single major events and large shifts, such as mass extinctions, tend to occur when there is a conjunction of events and processes, creating an extremely stressed environment. The single major event may be the final driving force to trigger a regime shift. If the event causes a regime shift, such as a mass loss of species, but there is no associated feedback change in the system, there is no threshold effect. It is, however, often difficult to determine whether feedback changes were involved. Examples such as this are included in the database in order to better understand the attributes of the system that distinguish between these kinds of big changes and those involving regime shifts with feedback changes. Class 2. No linkage, internally driven change in the ecological or social systemsCategory 2a: Internally driven shift in the ecological system with no external pressure from society or from environmental drivers outside the ecosystemA shift in this category may include either physical/chemical changes in the physical part of the ecosystem, or a shift involving biophysical interactions. A biological community can alter its environment (e.g., increasing water input or lowering soil pH), making it more suitable for a different community to develop (Wilson and Agnew 1992). For example, in mountainous areas where rainfall is marginal for forest development, fog precipitation intercepted by vegetation can increase water input to the system, enabling the growth of a taller, and sometimes different, vegetation type (Vogelmann et al. 1968). This in turn further increases the amount of fog precipitation available to the system, enabling a further change in vegetation. This positive feedback mechanism can result in a sharp boundary between adjacent vegetation types, with the forest type dependent on a threshold level of fog precipitation. A further example of an internally driven regime shift occurs in semi-arid regions, where mosaics of bare ground and bands or arcs of dense vegetation (originally called “brousse tigre” in West Africa) develop as a consequence of a threshold effect of water availability (Tongway and Ludwig 1990). On average, there is insufficient rainfall to maintain a continuous vegetative cover and, initially, rainfall runs off across mostly bare soil. Any slight impediment to the flow (a fallen log, a pile of litter) that allows the water to accumulate can result in a level of water infiltration above a threshold that allows significant growth to be maintained. The effect is reinforced over time, leading to a marked, banded landscape pattern, representing a shift to a spatially patterned ecosystem regime. van de Koppel and Rietkerk (2004) modelled this spatial patterning and identified feedback processes operating on multiple scales. Spatial concentration of water within vegetated patches invokes a strong positive feedback to plant growth at a local scale, and a negative feedback operates by depleting water at a larger scale. Class 3: Linked social–ecological systems, with a threshold change in only one systemCategory 3a: Shift in the ecological system, driven by the social systemGiven the profound impact of people on the world’s ecosystems, most of the examples in the database so far fall into this category. In these examples, the ecosystem has been altered, but this has not yet fed back to produce a reciprocal shift in the social system. We have used the database to identify three primary ways that people have impacted on the environment to effect shifts in ecosystem state: intensity of use, conversion of land cover, and addition of pollutants or new species. (i) Intensity Vegetation shift with grazing intensity—Regime shifts in savannas and rangelands have been attributed to grazing intensity, often interacting with fire. In rangeland livestock-production systems in semi-arid Australia (Fernandez et al. 2002, Anderies et al. 2002, Walker 1981), Argentina (Bisigato and Bertiller 1997), and Africa (Walker 1981), a threshold density of grazing animals has been implicated in switching vegetation type from “desirable” grassy to “undesirable” wooded systems or bare soil. In a switch to bare soil, heavy grazing removes the vegetation and a soil crust develops such that water can no longer penetrate the soil and perennial seeds can no longer germinate (Fernandez et al. 2002). A three-state system has been found in southeastern Australia (Anderies et al. 2002) in response to the interactions between vegetative growth, vegetation composition, and fire dynamics under varying levels of grazing pressure. A combination of social factors (livestock prices) and ecological conditions (effect of shrubs on grass production) determines the regime into which the system develops. Shifts in vegetation state with grazing intensity can also occur in wildlife reserves. The populations of herbivores in northern Botswana (Walker 1989) and Tanzania (Dublin et al. 1990) have fluctuated with levels of rainfall and rinderpest disease and the frequency and intensity of fires. The vegetation has alternated between grass dominance at times of high herbivore density and woody dominance at times of low herbivore density. Where park managers maintained the populations of herbivores at high densities (by providing artificial water supplies for the animals), regeneration of woody plants was prevented and the system remained in a grassy state. Cascading effects of harvesting—Harvesting of plants and animals can lead to a regime shift with a threshold population density below which competition or predation prevents the population from recovering. Significantly reducing or eliminating the top predator from a system can have a cascading effect at other levels, affecting ratios of species and leading to dominance of some other (“undesirable”) species, with the top predator unable to regain its former numbers. Examples include the interactions between sea otters, sea urchins, and kelp (Simenstad et al. 1978, Estes and Duggins 1995); fish, sea urchins, coral, and algae (Hughes 1994, Nystrom et al. 2000); and walleye and minnow (Walters and Kitchell 2001). In the latter example, walleye (Strizostedion vitrium) feed on minnow, which in turn feed on walleye larvae. It is suggested that heavy fishing of walleye adults has allowed minnow to become the key predator (Walters and Kitchell 2001). There is a threshold body size of walleye larvae above which minnows can no longer prey on them. The relative densities of minnow and walleye larvae below the predation threshold prevents recovery of the adult walleye population to former levels. (ii) Conversion Vegetation / land cover / climate change—Clearing of native vegetation over large areas (usually for agriculture) can lead to changes in climate variables (net surface radiation, relative humidity, temperature) that prevent a return to the original vegetation type. At large scales, several regions of the world exhibit regime shifts in the coupled atmosphere–biosphere system, the state dependent on the vegetation type and density (Higgins et al. 2002). In West Africa, the strength of the tropical monsoon influences the distribution of vegetation, but also depends upon that vegetation. Extensive logging may have altered the net surface radiation and entropy flux, reducing regional rainfall, with a possible collapse of the monsoon system (Zheng and Eltahir 1997, 1998). Similarly, in the Amazon Basin, deforested areas that now support shallower rooted vegetation, experience warmer, drier, and longer dry periods than the forested areas (Kleidon and Heimann 1999). The threshold in relation to rooting depth is linked to the amount of water transpired by the plants and relative atmospheric humidity. In parts of West Africa and the Amazon, the changed climate may have made it impossible for the vegetation to return to the original forest type because the original forests were established during a wetter period in Earth’s history. Effects on soil quality—Converting natural ecosystems into agricultural systems is the most widespread of all land-use changes. On several continents, soil salinity has been implicated in the collapse of agricultural systems, with possibly the first example being irrigated agricultural lands in ancient Mesopotamia (Tainter 2000). Clearing deep-rooted perennial vegetation leads to reduced transpiration of water from the soil. The excess water flows through the soil to the groundwater, resulting in a rise in the water table. In many of the world’s old (non-glaciated) soils, small amounts of salt dissolved in rainfall have accumulated in the soil profile, in a layer below the depth to which normal annual rainfall penetrates. In eastern Australia, once the water table has risen to approximately 2 m below the surface, the groundwater moves by capillary action through fine pores to the surface, carrying the salt with it. Agricultural production drops sharply (Talsma 1963, Gardner 1958, Murray-Darling Basin Ministerial Council 1999) and the salt remains in the soil for a long time, even if the water table drops. Habitat fragmentation—The degree of isolation of remnant patches in cleared landscapes, and the ability of the biota to disperse between these remnants can set up thresholds of population viability. Regime shifts have been observed in regard to population density with fragment size and isolation (Andren 1996), population persistence with habitat size and quality (Bascompte and Solé 1996, With and Crist 1995) and habitat quality with patch size and connectivity (Metzger and Decamps 1997, Bascompte and Solé 1996, With and Crist 1995). Two empirical studies, on lizards in Australia (Sarre et al. 1995, Wiegand et al. 2001) and butterflies in Finland (Hanski et al. 1995), demonstrate that thresholds of population viability are reached such that, below a critical minimum level of a habitat resource, the probability of population persistence falls quickly toward zero. (iii) Additions Eutrophication—We have encountered more examples of shifts in lake and ocean turbidity and eutrophication than of any other threshold process. Examples are available from several continents, in particular Europe (Meijer and Hosper 1997, Scheffer et al. 1997, Sondergaard et al. 1999) and North America (Carpenter et al. 1999, Larsen et al. 1981, Ludsin et al. 2001), where many shallow lakes, fed from streams in surrounding agricultural land, are now eutrophic. Many empirical studies make comparisons both temporally and spatially within and between lakes. Phosphorus and nitrogen entering the water from surrounding agricultural lands are the primary threshold variables, creating a shift from clear to turbid and eutrophic conditions in lakes. In some instances, the change of state is reversible, either through seasonal dilution or through human intervention—removing the nutrient source and/or manipulating the lake biota to encourage re-establishment of the macrophytic flora. Reversibility of the shift tends to be more likely in shallow than in deep lakes. Meijer (2000) identified a threshold level of total phosphorus concentration (approx. 0.15 mg P/L) where the system shifted from a clear to eutrophic state. Recovery showed a hysteretic effect, only beginning when the total phosphorus concentration dropped below approximately 0.10 mg P/L. Biological invasions—Non-indigenous plant and animal species now make up a large proportion of flora and fauna in many parts of the world. Extinctions of native species have been attributed to such invasions (e.g., introduced cats and European foxes in Australia (Burrows et al. 2003)), yet few examples have identified a threshold effect. One example of an invasive plant species is the introduced plant Myrica faya that invades young volcanic sites in Hawaii, where the growth of native plants is limited by naturally low nitrogen levels (Vitousek et al. 1987). Myrica faya has a nitrogen-fixing symbiont that quadruples the amount of nitrogen entering the soil. This leads to a change in plant species composition and possibly an irreversible shift in vegetation type in favor of “weedy” species (Walker and Vitousek 1991). Category 3b: Shift in the ecological system, causing a change in the social systemThere is ample evidence that ecological changes influence society, without (necessarily) causing a regime shift in the social system. Whether or not there is then a feedback effect from the social system, however, requires more careful consideration. We use the example (cited earlier) describing the effects of reversal of climate oscillations in the Pacific Ocean on populations of ocean flora and fauna (Hare and Mantua 2000). The regime shift was in the coupled climate–ocean system with a consequent, associated change in the biota. This biological regime shift then affected society, forcing an adjustment of fishing quotas and practices. It is, therefore, an example of a regime shift in an ecosystem influencing the social system. The paper does not suggest that the changes in the social system constituted a regime shift. Furthermore, although the effect on the social system (changed fishing practices) must have fed back to some extent to the ecosystem (fish populations) they did not feed back to the physical climate–ocean “system” that had undergone the original regime shift. Therefore, we include it as an example in this category. However, it serves to highlight that a tightly defined and clearcut categorization of effects, regime shifts, and feedbacks is difficult, and the classification procedure should not become a hindrance. The categories are used only as an aid for presentation, and to assist database searches. Category 3c: Shift in the social system, driven by the ecological systemIn this category, a change in ecosystem condition (but not necessarily a regime shift) results in a regime shift in the social system. These regime shifts often take the form of changes in rule-based management, where the adoption of a different set of rules governs the use of an ecological resource. Each rule set is a different regime in the social system. As the supply of an ecological resource changes, a different rule set (a different social regime) is imposed to regulate the use of that resource, aimed at preventing the ecosystem from shifting to a state that is deemed to be undesirable. The different rule set—usually management practices—is imposed once a defined level (or threshold) along an environmental gradient is passed. The “rules” may be in the form of norms (mutually agreed resource use) or may be enforceable by law, with penalties imposed for breaking the rules (Ostrom 1990). An example of such alternate rule regimes is an irrigated agricultural system in Valencia, Spain, where a three-state system of water allocation to farmers, dependent on water availability, has been operating for over 600 years (Ostrom 1990). When water is abundant, there is unrestricted water use; seasonally low water permits water supply to all, but under a rotation scheme for a fixed amount of time; and, in extreme drought, priority is given to farms most in need. These rules are enforceable, with fines imposed for infractions. Class 4: Linked social–ecological systems with reciprocal influences, but a shift in only one systemCategory 4a: Shift in the ecological system; society changes the state of the ecological system, which has a feedback effect on social behaviorTikopia in the Solomon Islands, is an example with several regime shifts in the ecological system (Kirch 1997). When the island was first settled around 900 B. C., Tikopia had tropical forest and nutrient-rich soils. For the first 1000 years of settlement the forest was burned and cleared for shifting cultivation. The soil was eroded and native flora and fauna were depleted, with several species becoming extinct. Instead of a collapse of both the human population and the environment, however, the feedback on social behavior led the society to change. They changed agricultural practices and developed an alternate system of multi-layered orchards that mimicked the structure of the former tropical forest. It could be argued that the change in society was a regime shift. However, our interpretation of the paper is that society altered its behavior, but this did not amount to a regime shift. The example nevertheless again illustrates that, based on published accounts of changes, deciding what is or is not a regime shift is not always clear cut. Category 4b: Shift in the social system; society uses the ecosystem, changes in which (often with an environmental shock) cause a regime shift in societySocieties may change in state (often collapse) from overusing an ecosystem, or from climatic changes that temporarily affect the ecosystem. Such shifts usually result from a complex set of events that lower the resilience of the society, making it unable to recover from an environmental shock (event). A prolonged drought of several decades may have been the final event that forced the collapse of the Chaco Canyon (Tainter 1988, Diamond 2001) and Maya (Tainter 1988, Gill 2000) societies. Both had established complex hierarchies that were supported largely by the farming population. This brought difficulties, especially in Chaco Canyon, where the climate was marginal for growing maize, the primary crop. A prolonged drought meant that diminishing returns could no longer fund the government hierarchies and infrastructure costs, resulting in a change in state (collapse) of the social system. Class 5: Linked social–ecological systems with reciprocal influences, shifts in both the ecological and social systemsEaster Island is a well-known example of regime shifts in both the ecosystem and society. Key species of native flora and fauna, crucial to human welfare, were decimated or eliminated through over-harvesting and the human population dropped from an estimated high of 10 000 to 2000 people by the time of European contact (Rainbird 2002). When it was first settled, around 800 A. D., the island was covered with tropical forest and had 43 species of breeding land and sea birds. The trees were felled for firewood, making gardens, building canoes, and rolling and levering the giant statues carved on the island. By 1600 A. D. the ecosystem had undergone a regime shift: all the trees and land birds, and all but one of the sea birds were extinct. Without trees, soil erosion became rife; canoes, needed for catching large fish and sea mammals for food, could no longer be built, which also made emigration impossible. The society shifted to a regime with a much-reduced population size and a behavioral change involving cannibalism.
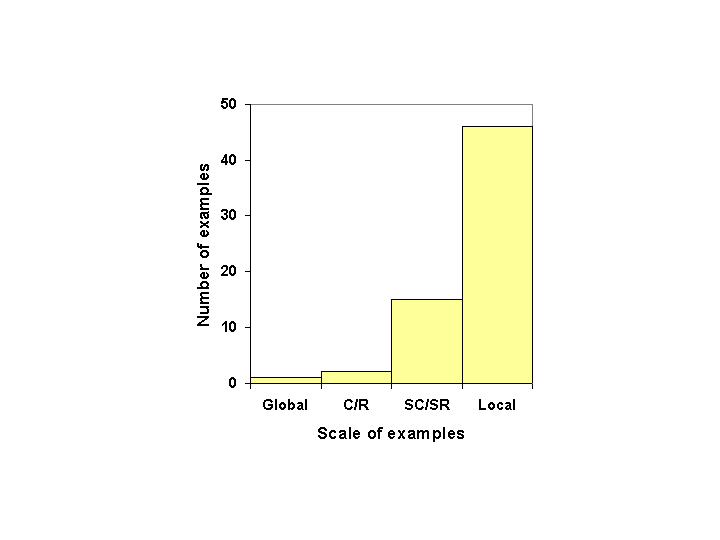
It is premature at this stage of the database development (64 examples) to attempt any in-depth analysis. We need substantially more examples before reliable conclusions can be drawn. However, to give some flavor of the sorts of insights we are seeking, the following are some initial observations arising from our considerations of the examples included so far. We have resisted the temptation to go beyond these few observations. DynamicsThresholds are not constant. Rather, the position of a threshold along a determining variable can change. In a three-regime, forest–cropland–grassland system in Southeast Asia (Trenbath et al. 1989, Conway 1997), the maximum level (threshold) of cropping possible before the land will no longer revert to forest depends on the prior management (including fertilization) of that land. The threshold time for a phase shift from grassland back to forest depends on whether the grassland was grazed or burned. In the rangelands example described by Walker (1993), if the grass layer consists of all perennials, the threshold ratio of shrubs (S) to grass (G) (the slow-changing, controlling variable) is higher than if the grass layer consists of only annuals. Resilience is a dynamic property and, using the metaphor of a basin of attraction, changes in thresholds raise the question of how to measure changes in the topology of the basin. Not only might the position of a threshold change, but so too might the depth of the basin, making it easier or harder to approach the threshold. Although it is clear that different systems vary with regard to their resistance to change, to date no examples in the database make mention of a change in this aspect of resilience. This raises the question of whether resistance does change and, if so, how significant such changes might be in relation to crossing thresholds. Perhaps their absence in published accounts is related to the difficulty in observing them. Nature of the Threshold and Possible Regime ShiftsOf the 64 examples in the database, 24 have undergone an irreversible regime shift, and 32 are reversible, with eight of these showing a hysteresis effect. In one example, the direction of the shift is conditional on the management practices in the initial state. In a further eight examples, reversibility is either unknown or not applicable to the example. It is premature to attempt an analysis of the system attributes that lead to this set of threshold types, but once the database has an adequate representation of system types, this should be a high priority. ScaleChanges in scale influence resilience (and, therefore, the positions of thresholds). In the example of the ancient Chaco Canyon (Tainter 1988, Diamond 2001) and Maya (Tainter 1988, Gill 2000) civilizations, increasing the scale (regionalization) initially increased resilience by adding a diversity of ecosystem types that would provide a variety of different crop types and other ecosystem uses, that responded differently to climate variations and, therefore, created a buffer against the effects of a variable climate. With continued expansion, ecosystems were merely replicated, but the proportional costs of maintaining the society increased. The increasing ratio of costs to production led to declining flexibility, and hence resilience. The threshold level of drought that precipitated a collapse in the society decreased. This interpretation of the Chaco Canyon system raises an appropriate question for all SESs that have undergone a change: was it a change in scale that led to the threshold being crossed? Threshold changes on a large scale (e.g., reversal of ocean currents) are more rare and difficult to measure. Figure 4 shows that, of the 64 examples in the database, most of the regime shifts are small in scale (landscape scale) and the number of examples gets progressively smaller as the scale becomes larger.
Kinds of Feedback Changes A characteristic feature of a threshold is a change in system feedbacks. We need more examples to derive a categorization of feedback effects involved in thresholds that will aid future research into SESs. However, four common kinds of feedback changes have been identified in the examples so far.
Examples of thresholds between alternate attractors (e.g., lakes and rangelands) make mention of external shocks that can push the system across the threshold between the alternate attractors. As resilience declines, the amount of disturbance needed to cross the threshold declines. In many of the published papers on thresholds, however, and particularly accounts of very large-scale shifts (warmer boreal forests to colder tundra along a gradient of declining boreal forest cover (Bonan et al. 1992), ocean circulation regime shifts (Higgins et al. 2002) and changes in vegetation and climate regimes in West Africa (Zheng and Eltahir 1997, 1998)), there is no mention of shocks. The impression gained is of a regime shift of type B in Fig. 1, with an inevitable change in regime as the system’s “slow” variable crosses the threshold, without the system having some intermediate range of the slow variable in which there are alternate attractors and in which external shocks and variability play an important role. This may well be due to inadequate information. It calls for care in using only one published account of a regime shift, without further investigating the nature of the threshold. Threshold Models, Management, and PerceptionsMany of the management and governance difficulties reported in the database examples are associated with having the wrong mental model about system thresholds. Either the “model” says there is no threshold where there is one, or there is a threshold where there is not. If the threshold has not previously been crossed, then the regime shift generally comes as a surprise. Did the community on Easter Island (Rainbird 2002) consider the long-term consequences as the last trees were being felled? A corollary of this observation is that a change in mental model, or a shift in rules and norms in the social system, often follows an undesirable change in the ecosystem. Alternatively, the right mental model may be established but is subsequently ignored by managers, perhaps for short-term political or economic reasons. Even though government authorities were alerted to the long-term consequences of increased salt levels at the soil surface as a result of clearing native vegetation in southwestern Western Australia (Beresford et al. 2001), this advice was ignored for many decades and land clearance was promoted for political and economic gain. The legacy today is large tracts of salinized land. Threshold ConsequencesThe consequences of crossing a threshold are context dependent. The threshold is sometimes known and the decision on what to do about it depends on the consequences of crossing it. In some rangelands, depending on soil type and rainfall (Anderies et al. 2002), the effects of shrubs on grass growth and on the costs of livestock mustering are such that it is economically worthwhile to make initial high profits from high stocking rates, allow the system to change into a shrubby state, and continue to make a living, albeit at a somewhat lower level. In other rangeland types, the phenomenon is the same (i.e., a shift from a grassy to a shrubby state), but the consequences are quite different, with the optimal economic strategy being to avoid the regime shift.
A key question in the dynamics of any system is whether or not that system has, or is likely to have, thresholds. Can a threshold be identified before it has been crossed (Carpenter 2003)? From the examples in the database, the question can sometimes be answered by extrapolation from similar closely related systems (e.g., in lakes). So far, we have found no published examples where a new kind of threshold has been predicted before it has been experienced. How to do this is an obvious question for any future research agenda on thresholds. Is a threshold an emergent property of some underlying set of attributes of a system? Is there a relationship, for example, between the network topology of a system and the likelihood of a threshold? Which research avenues will allow us to best approach this question? Research on a typology of thresholds is a priority topic in the emerging area of “sustainability science,” and it requires a rich database of empirical data. The database currently reflects the eclectic nature of our interests and contacts. More examples are required in all categories to make it an effective and useful resource tool. We welcome further examples in all five classes, especially where there are empirical data to support the described regime shift.
Responses to this article are invited. If accepted for publication, your response will be hyperlinked to the article. To submit a comment, follow this link. To read comments already accepted, follow this link.
Anderies, J. M., M. A. Janssen, and B. H. Walker. 2002. Grazing management, resilience, and the dynamics of a fire-driven rangeland system. Ecosystems 5(1):23–44. Andren, H. 1996. Population responses to habitat fragmentation: statistical power and the random sample hypothesis. Oikos 76(2):235–242. Bascompte, J. and R. V. Solé. 1996. Habitat fragmentation and extinction thresholds in spatially explicit models. Journal of Animal Ecology 65(4):465–73. Beisner, B. E., D. T. Haydon, and K. Cuddington. 2003. Alternative stable states in ecology. Frontiers in Ecology and the Environment 1(7):376–82. Beresford, Q., H. Bekle, H. Phillips, and J. Mulcock. 2001. The salinity crisis: landscapes, communities and politics. University of Western Australia Press, Crawley, Western Australia. Bisigato, A. J., and M. B. Bertiller. 1997. Grazing effects on patchy dryland vegetation in northern Patagonia. Journal of Arid Environments 36(4):639–53. Bonan, G. B., D. Pollard, and S. L. Thompson. 1992. Effects of boreal forest vegetation on global climate. Nature 359:716–18. Burrows, N. D., D. Algar, A. D. Robinson, J. Singara, B. Ward, and G. Liddelow. 2003. Controlling introduced predators in the Gibson Desert of Western Australia. Journal of Arid Environments 55(4):691–713. Carpenter, S. R. 2003. Regime shifts in lake ecosystems: pattern and variation. in O. Kinne, editor. Excellence in Ecology: 15. International Ecology Institute, Germany. Carpenter, S. R., D. Ludwig, and W. A. Brock. 1999. Management of eutrophication for lakes subject to potentially irreversible change. Ecological Applications 9(3):751–71. Conway, G. 1997. The doubly green revolution: food for all in the twenty-first century. Comstock Publishing Associates, Ithaca, New York, USA. Diamond, J. 2001. Archaeology—tree trail to Chaco Canyon. Nature 413:687–90. Dublin, H. T., A. R. E. Sinclair, and J. McGlade. 1990. Elephants and fire as causes of multiple stable states in the Serengeti-Mara woodlands. Journal of Animal Ecology 59:1147–64. Estes, J. A., and D. O. Duggins. 1995. Sea otters and kelp forests in Alaska—generality and variation in a community ecological paradigm. Ecological Monographs 65(1):75-100. Fernandez, R. J., E. R. M. Archer, A. J. Ash, H. Dowlatabadi, P. H. Y. Hiernaux, J. F. Reynolds, C. H. Vogel, B. H. Walker, and T. Wiegand. 2002. Degradation and recovery in socio-ecological systems. Pages 297–323 in F. Reynolds, and D. M. Stafford Smith, editors. Global desertification: do humans cause deserts? Dahlem University Press, Berlin, Germany. Frankham, R. 1995. Inbreeding and extinction—a threshold effect. Conservation Biology 9(4):792–799. Gardner, W. R. 1958. Some steady state solutions of the unsaturated moisture flow equation with application to evaporation from a watertable. Soil Science 85:228–232. Gill, R. B. 2000. The great Maya droughts: water, life and death. University of New Mexico Press, Albuquerque, New Mexico, USA. Gunderson, L. H. 2001. Managing surprising ecosystems in southern Florida. Ecological Economics 37(3):371–378. Hanski, I., J. Poyry, T. Pakkala, and M. Kuussaari. 1995. Multiple equilibria in metapopulation dynamics. Nature 377:618–621. Hare, S. R., and N. J. Mantua. 2000. Empirical evidence for North Pacific regime shifts in 1977 and 1989. Progress in Oceanography 47(2-4):103–145. Higgins, P. A. T., M. D. Mastrandrea, and S. H. Schneider. 2002. Dynamics of climate and ecosystem coupling: abrupt changes and multiple equilibria. Philosophical Transactions of the Royal Society of London Series B-Biological Sciences 357:647–655. Holling, C. S. 1973. Resilience and stability of ecological systems. Annual Review of Ecological Systems 4:1–23. Hughes, T. P. 1994. Catastrophes, phase shifts, and large-scale degradation of a Caribbean coral reef. Science 265:1547–1551. Kirch, P. V. 1997. Microcosmic histories: island perspectives on “global” change. American Anthropologist 99(1):30–42. Kleidon, A., and M. Heimann. 1999. Deep-rooted vegetation, Amazonian deforestation, and climate: results from a modelling study. Global Ecology and Biogeography 8(5):397–405. Larsen, D. P., D. W. Schultz, and K. W. Malueg. 1981. Summer internal phosphorus supplies in Shagawa Lake, Minnesota. Limnology and Oceanography 26(4):740–753. Ludsin, S. A., M. W. Kershner, K. A. Blocksom, R. L. Knight, and R. A. Stein. 2001. Life after death in Lake Erie: nutrient controls drive fish species richness, rehabilitation. Ecological Applications 11(3):731–746. Ludwig, D., Walker, B. and C. S. Holling. 1997. Sustainability, stability, and resilience. Conservation Ecology 1(1):7. [Online.] URL: http://www.consecol.org/vol1/iss1/art7. May, R. M. 1977. Thresholds and breakpoints in ecosystems with a multiplicity of stable states. Nature 269:471–477. Meijer, M.-L. 2000. Biomanipulation in the Netherlands: 15 years of experience. Ministry of Transport, Public Works and Water Management, Institute for Inland Water Management and Wastewater Treatment (RIZA), Lelystad, the Netherlands. Meijer, M. L., and H. Hosper. 1997. Effects of biomanipulation in the large and shallow Lake Wolderwijd, the Netherlands. Hydrobiologia 342:335–349. Metzger, J. P., and H. Decamps. 1997. The structural connectivity threshold: an hypothesis in conservation biology at the landscape scale. Acta Oecologica-International Journal of Ecology 18(1):1–12. Moss, B., J. Stansfield, K. Irvine, M. Perrow, and G. Phillips. 1996. Progressive restoration of a shallow lake: a 12-year experiment in isolation, sediment removal and biomanipulation. Journal of Applied Ecology 33(1):71–86. Muradian, R. 2001. Ecological thresholds: a survey. Ecological Economics 38:7–24. Murray-Darling Basin Ministerial Council. 1999. The salinity audit of the Murray-Darling Basin. Murray-Darling Basin Commission, Canberra, Australia. [Online.] URL: http://www.mdbc.gov.au/naturalresources/pdf/Final_Salt_Audit.pdf Nystrom, M., C. Folke, and F. Moberg. 2000. Coral reef disturbance and resilience in a human-dominated environment. Trends in Ecology and Evolution 15(10):413–417. Ostrom, E. 1990. Governing the commons: the evolution of institutions for collective action. Cambridge University Press, Cambridge, UK. Pech, R. P., A. R. E. Sinclair, A. E. Newsome, and P. C. Catling. 1992. Limits to predator regulation of rabbits in Australia—evidence from predator-removal experiments. Oecologia 89(1):102–112. Rainbird, P. 2002. A message for our future? The Rapa Nui (Easter Island) ecodisaster and Pacific island environments. World Archaeology 33(3):436–451. Robblee, M. B., T. R. Barber, P. R. Carlson, M. J. Durako, J. W. Fourqurean, L. K. Muehlstein, D. Porter, L. A. Yarbro, R. T. Zieman, and J. C. Zieman. 1991. Mass mortality of the tropical seagrass Thalassia testudinum in Florida Bay (USA). Marine Ecology-Progress Series 71(3):297–299. Sarre, S., G. T. Smith, and J. A. Meyers. 1995. Persistence of two species of gecko (Oedura reticulata and Gehyra variegata) in remnant habitat. Biological Conservation 71(1):25–33. Scheffer, M., and S. R. Carpenter. 2003. Catastrophic regime shifts in ecosystems: linking theory to observation. Trends in Ecology and Evolution 18(12): 648–656. Scheffer, M., S. Carpenter, J. A. Foley, C. Folke, and B. Walker. 2001. Catastrophic shifts in ecosystems. Nature 413:591–596. Scheffer, M., S. Rinaldi, A. Gragnani, L. R. Mur, and E. H. Vannes. 1997. On the dominance of filamentous cyanobacteria in shallow, turbid lakes. Ecology 78(1):272–282. Scheffer, M., S. Szabo, A. Gragnani, E. H. van Nes, S. Rinaldi, N. Kautsky, J. Norberg, R. M. M. Roijackers, and R. J. M. Franken. 2003. Floating plant dominance as a stable state. Proceedings of the National Academy of Sciences of the United States of America 100(7):4040–4045. Simenstad, C., J. Estes, and K. Kenyon. 1978. Aleuts, sea otters, and alternative stable-state communities. Science 200: 403–411. Sondergaard, M., J. P. Jensen, and E. Jeppesen. 1999. Internal phosphorus loading in shallow Danish lakes. Hydrobiologia 409: 145–152. Tainter, J. A. 1988. The collapse of complex societies. New studies in archaeology, Cambridge University Press, Cambridge, UK. Tainter, J. A. 2000. Problem solving: complexity, history, sustainability. Population and Environment 22(1): 3–41. Talsma, T. 1963. The control of saline groundwater. Mededelingen van de Landbouwhogeschool Wageningen - Nederland 63(10):1–68. Tongway, D. J., and J. A. Ludwig. 1990. Vegetation and soil patterning in semi-arid mulga lands of eastern Australia. Australian Journal of Ecology 15:23–34. Trenbath, B. R., G. R. Conway, and I. A. Craig. 1989. Threats to sustainability in intensified agricultural systems: analysis and implications for management. Pages 337–365 in S. R. Gliessman, editor. Agroecology: researching the ecological basis for sustainable agriculture. Springer-Verlag, New York, New York, USA. van de Koppel, J., and M. Rietkerk . 2004. Spatial interactions and resilience in arid ecosystems. American Naturalist 163(1):113–121. Vitousek, P. M., L. R. Walker, L. D. Whiteaker, D. Muellerdombois, and P. A. Matson. 1987. Biological invasion by Myrica faya alters ecosystem development in Hawaii. Science 238:802–804. Vogelmann, H. W., T. Siccama, D. Leedy, and D. C. Ovitt. 1968. Precipitation from fog moisture in the Green Mountains of Vermont. Ecology 49:1205–1207. Walker, B. H. 1981. Stability properties of semiarid savannas in southern African game reserves. Pages 57–67 in P. A. Jewell, S. Holt, and D. Hart, editors. Problems in management of locally abundant wild mammals. Academic Press Inc. Walker, B. 1989. Diversity and stability in ecosystem conservation. Pages 121–130 in D. Western and M. Pearl, editors. Conservation for the twenty-first century. Oxford University Press, London, UK. Walker, B. H. 1993. Rangeland ecology—understanding and managing change. Ambio 22(2–3):80–87. Walker, L. R., and P. M. Vitousek. 1991. An invader alters germination and growth of a native dominant tree in Hawaii. Ecology 72(4):1449–1455. Walters, C., and J. F. Kitchell. 2001. Cultivation/depensation effects on juvenile survival and recruitment: implications for the theory of fishing. Canadian Journal of Fisheries and Aquatic Sciences 58(1):39–50. Wang, G. L., and E. A. B. Eltahir. 2000. Role of vegetation dynamics in enhancing the low-frequency variability of the Sahel rainfall. Water Resources Research 36(4):1013–1021. Wiegand, K., S. D. Sarre, K. Henle, T. Stephan, C. Wissel, and R. Brandl. 2001. Demographic stochasticity does not predict persistence of gecko populations. Ecological Applications 11(6): 1738–1749. Wilson, J. B., and A. D. Q. Agnew. 1992. Positive-feedback switches in plant-communities. Advances in Ecological Research 23:263–336. With, K. A., and T. O. Crist. 1995. Critical thresholds in species responses to landscape structure. Ecology 76(8): 2446–2459. Zheng, X. Y. and E. A. B. Eltahir. 1997. The response to deforestation and desertification in a model of West African monsoons. Geophysical Research Letters 24(2):155–158. Zheng, X. Y. and E. A. B. Eltahir. 1998. The role of vegetation in the dynamics of West African monsoons. Journal of Climate 11(8):2078–2096. Zinsmeister, W. J. 1998. Discovery of fish mortality horizon at the K–T boundary on Seymour Island: re-evaluation of events at the end of the Cretaceous. Journal of Paleontology 72(3):556–571.
Address of Correspondent: Brian Walker CSIRO Sustainable Ecosystems, PO Box 284, Canberra, ACT 2601, Australia Phone: 61-2-62421740 Fax: 61-2-62421677 Brian.Walker@csiro.au
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||
| Home | Archives | About | Login | Submissions | Notify | Contact | Search | |||||||||||||||||||||||||||||||||||||||||||||||||||||