|
|
|
Copyright © 2004 by the author(s). Published here under license by The Resilience Alliance.
Go to the pdf version of this article.
The following is the established format for referencing this article:
Lusseau, D. 2004. The hidden cost of tourism: detecting long-term effects of tourism using behavioral information. Ecology and Society 9(1): 2. [online] URL: http://www.ecologyandsociety.org/vol9/iss1/art2/
A version of this article in which text, figures, tables, and appendices are separate files may be found by following this link.
Report, part of Special Feature on Ralf Yorque Memorial Competition 2004 The Hidden Cost of Tourism: Detecting Long-term Effects of Tourism Using Behavioral Information David Lusseau
University of Otago
- Abstract
- Introduction
- Methods
- Results
- Discussion
- Responses to this Article
- Acknowledgments
- Literature Cited
Increasingly, whales and dolphins are the focus of tourism activities in many coastal locations. Although these activities can affect individuals and populations of cetaceans, the biological significance and hence the cost of these impacts are as yet largely unknown. This study assessed the effects of boat interactions on the behavioral budget of two populations of bottlenose dolphins (Tersiops truncatus) living in similar fjords but exposed to different levels of tourism activities. This comparison makes it possible to assess the costs of short-term avoidance strategies and the threshold at which those strategies are no longer effective. The effects of boat interactions were the same in both fjords. The resting state was the most sensitive to interactions; socializing was less sensitive. Short-term displacement was a typical response to boat exposure: dolphins were more likely to travel after an interaction with a vessel. Although the behavioral budgets of these populations were significantly altered during interactions with boats, their overall behavioral budgets were unchanged. Dolphins in Milford Sound actively avoided boat interactions, possibly to maintain their overall behavioral budget unchanged. This active avoidance led to avoidance of the area. Characteristics of dolphin-boat interactions in Milford Sound suggest that the advantages gained by short-term avoidance are lost if, on average, fewer than 68 min elapse between successive interactions with boats. If dolphin-boat interactions were more frequent than this, the dolphins switched to a longer-term response: area avoidance.
KEY WORDS: Tursiops truncatus, Markov chain modeling, avoidance, behavioral budget, boat-dolphin interactions, boats, bottlenose dolphin, impact assessment, long-term effects, tourism, whale-watching.
Published: January 12, 2004
Attempts to assess the effects of tourism activities on cetaceans such as whales and dolphins have been the focus of considerable research effort over the last decade. Several studies have detected horizontal avoidance, increased dive intervals, increased speed, and variations in vocalizations (Kruse 1991, Corkeron 1995, Janik and Thompson 1996, Bejder et al. 1999, Nowacek et al. 2001, Van Parijs and Corkeron 2001, Williams et al. 2002). However, the biological importance of these changes still needs to be established. Changes in behavioral budgets, assessed using behavioral states, can provide information on the biological significance of an impact. The behavioral budget is directly related to the energy budget of individuals and populations. This parameter has so far not been explored because of difficulties in sampling and analysis (Mann 2000, Lusseau 2003a) that are especially important in the context of detecting impacts. The technique described here provides a way to assess the impacts of tourism activities on behavioral budgets.
Because behavioral states are mutually exclusive categories arbitrarily defined along the behavior continuum by an observer, they can be thought of as mutually dependent, and therefore their sequencing in time can be assessed using Markov chains. Stochastic processes such as Markov chains can be used to measure with precision the effects of different factors on a dataset, and are consequently valuable tools for ecological impact assessment (Tanner et al. 1994, Hill and Caswell 2001). I have shown how Markov chain modeling can be used to detect the impact of boat interactions on the behavioral budget of dolphins (Lusseau 2003a). It is possible to detect variations in responses to boat interactions in different populations by comparing their control and impact Markov chains. This study used that approach to compare the effects of boat interactions in two populations of bottlenose dolphins (Tursiops truncatus) exposed to different levels of boat traffic. These two populations live in adjacent and similar environments (Fig. 1). As a result, they come as close as possible to being true ecological replicates, and I therefore assumed that the differences in behavioral budgets were mainly attributable to the levels of boat traffic.
|
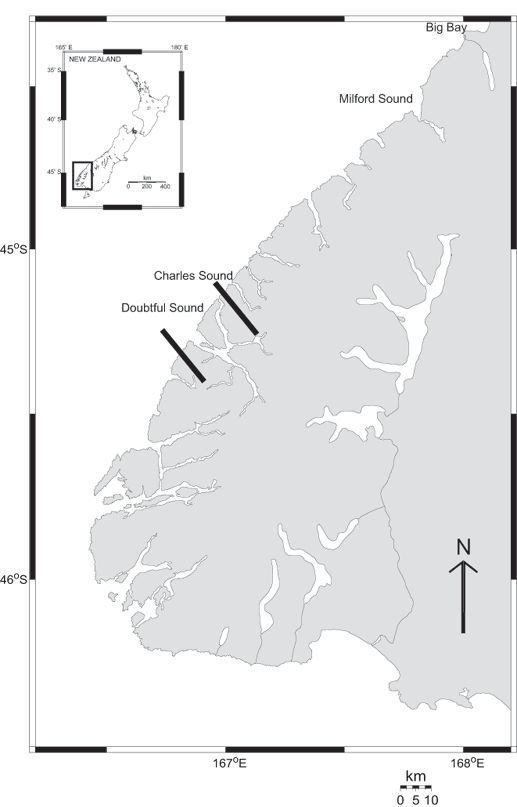
Fig. 1. Location of the two study sites. The home range of the Milford Sound population spans all fjords, bays, and the lake between Big Bay and Charles Sound. The Doubtful Sound population has not been observed outside the fjords located between the two black bars, i.e., the Doubtful/Thompson/Bradshaw fjord complex.
|
The Milford Sound population spans several fjords, bays, and a lake from Charles Sound to Big Bay (Fig. 1) and numbers approximately 50 individuals (Lusseau et al. 2002). Despite very extensive study, the Doubtful Sound population has not been observed outside the Doubtful/Thompson/Bradshaw fjord complex (Fig. 1) during the past decade (Lusseau et al. 2003). It is composed of 65 individuals (Lusseau et al. 2003). Both populations have a high mark rate (> 90%), which means that more than 90% of all individuals in both populations can be easily recognized (Williams et al. 1993, Lusseau et al. 2003). No interaction between the two populations, or swapping of members, was observed during this period despite several thousands of hours spent looking for dolphins in both locations (Lusseau et al. 2002, Lusseau et al. 2003). It is therefore safe to assume that these populations are to a large extent isolated. Both are located at the southern extreme of the species' range. Tourism activities are well established in Milford Sound, with more than 17,000 boat trips undertaken during this study, i.e., between June 2000 and May 2002. In contrast, 3500 boat trips were offered in Doubtful Sound during this study (Lusseau et al. 2002). Because of the isolation of these fjords, tourism activities account for most of the boat traffic; fishing vessels and private recreational vessels make up less than 5% (Lusseau 2002).
The goal of this study was to determine how interactions with tour boats affected the temporal dynamics of dolphin behavioral states and how these effects impacted the behavioral budget of bottlenose dolphins in Doubtful Sound and Milford Sound. This study provides a window into the energetic cost of these responses, and therefore their biological significance.
I conducted systematic surveys of the fjords from June 2000 to May 2002. Once a school was detected, the identities of the individuals in the school were recorded via digital video. The behavioral state of the school was sampled every 15 min, and the principal behavioral state (Table 1) of the school was categorized via scan sampling (Altmann 1974). Behavioral states were defined to be mutually exclusive and cumulatively inclusive; as a whole, they described the entire behavioral budget of the dolphins. These states were similar to the ones used in other studies (Shane 1990, Bearzi et al. 1997). I sampled focal schools to understand the effect of boat interactions at the school level, rather than at the individual level. Moreover, dolphins live in large schools in Fiordland, with an average of 17 individuals in those of Doubtful Sound. It was therefore more appropriate to follow focal groups (Mann 2000). Scan sampling of individuals within the school was used instead of focal group sampling because of the observer bias inherent in the latter (Mann 2000). Observations ended when the weather deteriorated, the focal school was lost because of changes in sighting conditions, or fading light made observations difficult. As a result, the end of a sequence of observations was not dependent on the behavior of the focal school. I used a 4.8-m aluminium boat powered by a 50-hp, four-stroke outboard engine to conduct these surveys. The presence of the observation platform did not appear to affect the behavior of focal schools (see Lusseau 2003a, b for more details). Generally, the observing vessel was kept parallel to the dolphin school and followed the school slightly from the rear. This position was not changed when other vessels interacted with the focal school.
|
Table 1. Definitions of the behavioral states.
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Developing Markov chains
Markov chains quantify the dependence of an event on preceding events. This dependence can be affected by any factor occurring between events. I therefore decided to assess the difference in transition from one event to another depending on the presence of boat interactions during that sequence and depending on the location. Because boat interactions lasted on average less than a behavioral sampling interval, I concentrated only on a first-order Markov chain model (see Lusseau 2003a for details of analysis). Moreover, first-order Markov chains proved to be a good approximation of the dataset in Doubtful Sound (Lusseau 2003a).
Data for each fjord were compiled into two two-way contingency tables of preceding behavioral state vs. succeeding behavioral state, depending on whether or not an interaction with a boat had occurred between the samples. Hence, one table represents potential impact situations, and the other is the control. These were compared to detect the effect of boat interactions.
Log-linear analysisA four-way contingency table was constructed by merging the impact and control chains from both locations (two locations, two boat presence states, five preceding behaviors, and five succeeding behaviors). A log-linear analysis was applied to assess the independence of the behavior transitions from boat presence and location and the interaction of boat presence and location. The following abbreviations are used in the description of the models: L for location, B for boat presence, P for preceding behavior, and S for succeeding behavior. The effects of both factors on behavior transitions were analyzed using the technique described by Caswell (2001).
The log-linear analysis fits a saturated model to the dataset, i.e., a model that considers all interactions, indicated by BLPS in this case. The effects of the different factors can be tested by comparing different fitted models using likelihood ratio tests. Here the null hypothesis is that succeeding behaviors are independent of boat presence and location, given preceding behaviors. This null hypothesis corresponds to the null model PS, PBL (Fig. 2).
|
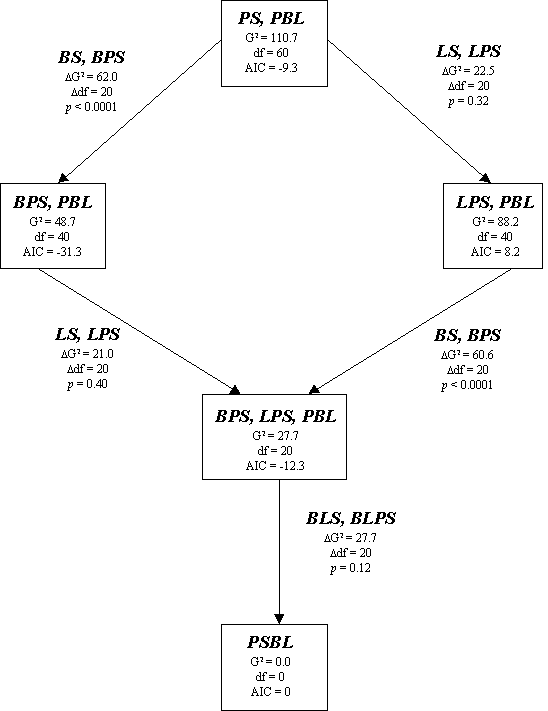
Fig. 2. Tests of boat presence (B) and location (L) effects on transitions between preceding behavior (P) and succeeding behavior (S). Models and their respective goodness-of-fit G2 statistics, degrees of freedom (df), and Akaike Information Critera (AIC) values are shown in the boxes. Terms added, with the corresponding increment in G2 and degrees of freedom, are shown along the arrows (adapted from Caswell 2001).
|
The effects of boat presence and location can each be tested by comparing different pairs of models as shown in Fig. 2. Beginning with the null model at the top of the figure, I add the boat presence effect by including the terms BS and BPS. Their effect is evaluated by comparing the null model and BPS, PBL because adding the terms BS and BPS to the null model results in the model BPS, PBL. The significance of the boat presence effect can then be tested by comparing the goodness-of-fit of both models. The difference in goodness-of-fit (ΔG2 = G2BPS,PBL - G2PS,PBL) is the likelihood ratio testing BS, BPS with degrees of freedom equal to the difference in the degrees of freedom for the two models. Similarly, the comparison of the null model to LPS, PBL tests for the effect of location (Fig. 2). The effect of location can also be tested by adding a location effect to the model already considering a boat presence term (BPS, LPS, PBL vs. BPS, PBL). It is also possible to test for a boat presence effect by adding this term to the model considering a location effect (BPS, LPS, PBL vs. LPS, PBL). Finally, the interaction of boat presence and location can be tested by comparing the saturated model and BPS, LPS, PBL.
The best-fitting model was selected using Akaike Information Criteria (AIC). AIC estimate the amount of information a model provides and penalize models for any excess parameters needed to explain the data (Anderson et al. 2000, Caswell 2001).
Impact assessmentTransition probabilities from preceding to succeeding behavior were then determined in both control and impact chains (Lusseau 2003a). Each transition is a proportion of the time that a succeeding behavior was observed following a preceding behavior. Therefore, the effect of boat interactions on the behavior-transition probability matrix was tested using a Z-test for proportions (Fleiss 1981).
To assess the effects of location and boat presence on the behavioral budget, budgets in each location and boat state were estimated from the left eigenvectors of the dominant eigenvalues of the transition matrices (see Caswell 2001 and Lusseau 2003a for more details). Eigenanalyses of the transition matrices were performed using PopTools 2.3, an Excel Add-in developed by CSIRO (http://www.cse.csiro.au/CDG/poptools). Differences between the four behavioral budgets (control vs. impact and Milford vs. Doubtful) were tested using the Z-test for proportions (Fleiss 1981), and 95% confidence intervals were calculated using Wilson's procedure as described in Newcombe (1998).
Forecast modelingTo understand the biological significance of the observed impacts, it is necessary to determine how the effects of boat interaction affect the overall behavioral budget of the populations. Boat-dolphin interactions have been described in both fjords (Lusseau 2002, Lusseau et al. 2002, Lusseau 2003c), so that the average amount of time dolphins spend with boats on a given day can be estimated, i.e., how long they spend in an "impact" behavioral budget. It is therefore possible to calculate a cumulative behavioral budget that takes into consideration the amount of time dolphins spend in a control behavioral budget and in an impact behavioral budget. It is consequently possible to detect whether this cumulative behavioral budget is significantly different from the control budget using a Z-ratio test for proportions (Fleiss 1981) for each behavioral state.
For example, if dolphins spend 12% of their time with boats during the study period, their cumulative budget is calculated as follows:
|
cumulative budget = (0.12 x impact budget) + (0.88 x control budget).
| (1) |
Assuming changes are linear, it is also possible to calculate at what level of boat interaction intensity the cumulative behavioral budget becomes different from the control budget by artificially varying the amount of time dolphins spend with boats from 0 to 100%.
Forecasting time needed to recover from interactions with boatsAs with any other impact, dolphins may require a certain amount of time after a boat interaction to recover the energetic cost involved in avoiding a stressful situation (Moberg 2000). If the time between two boat interactions is less than the necessary recovery period, the cost of interactions will cumulate (Moberg 2000). If this cumulated cost reaches a biologically significant level, dolphins have to alter their movement/behavior to maximize periods between two boat interactions. Dolphin-boat interactions would therefore no longer be random and would depend on the number of boats present on the water. I therefore assessed the relationship between boat traffic, measured as the number of trips offered per season, and the average time elapsed between two boat interactions each season in both fjords. For each season, I constructed a first-order Markov chain to assess the likelihood that dolphins would go from a behavior transition without interaction to a transition without interaction. From this probability, I approximated the average length of time without boat interaction from the mean of the geometric distribution characterized by p(no interaction-no interaction) (Guttorp 1995):
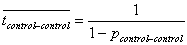 , , | (2) |
with a standard error of:
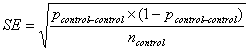 . . | (3) |
The length of time without a boat interaction corresponds here to the length of time the dolphins stayed in a control behavioral budget (pno interaction-no interaction = pcontrol-control). By the same token, ncontrol is the number of transitions that start with a control situation.
During the study period, I spent 137 days (879.2 h) looking for dolphins in Doubtful Sound and 112 days (505.5 h) in Milford Sound. I spent 716.5 h (more than 133 days) following focal groups in Doubtful Sound and 296.1 h (more than 53 days) in Milford Sound. Dolphins spent 10.8 and 12.8% of the time I spent observing them interacting with boats in Doubtful and Milford Sounds, for totals of 373 and 248 interactions, respectively (Lusseau et al. 2002). Despite this similar level of interaction, there were roughly five times more boat trips undertaken in Milford Sound than in Doubtful Sound (Lusseau et al. 2002).
Over the study period, I observed 1782 behavior transitions, i.e., passage from a preceding to a succeeding behavior, in Doubtful Sound and 532 transitions in Milford Sound. There were 375 and 172 impact transitions, respectively, in Doubtful and Milford Sounds. As seen in Table 2, all first-order Markov chains provided more information than zero-order chains (Guttorp 1995, Lusseau 2003a). The behavioral state of focal schools depended on the state they were in 15 min previously.
|
Table 2. Chain order selection using Bayes Information Criterion (BIC). A higher-order chain provides more information than a lower-order one if ΔBIC > 9.2 (Guttorp 1995).
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Log-linear analysis
The saturated model did not provide a significant improvement over the BPS, LPS, PBL model, indicating that there was no interaction between the location and boat presence terms (Fig. 2). Location did not have a significant effect on behavior transitions (Fig. 2). In both tests, the LS, LPS effect was not significant. The effect of boat presence was highly significant (Fig. 2). The Akaike Information Criteria difference between the best model (BPS, PBL) and the next best model (BPS, LPS, PBL) was overwhelming (16.8). Consequently, BPS, PBL was the best-fitting model by far. Boat presence significantly affected behavior transitions in the same fashion in both fjords.
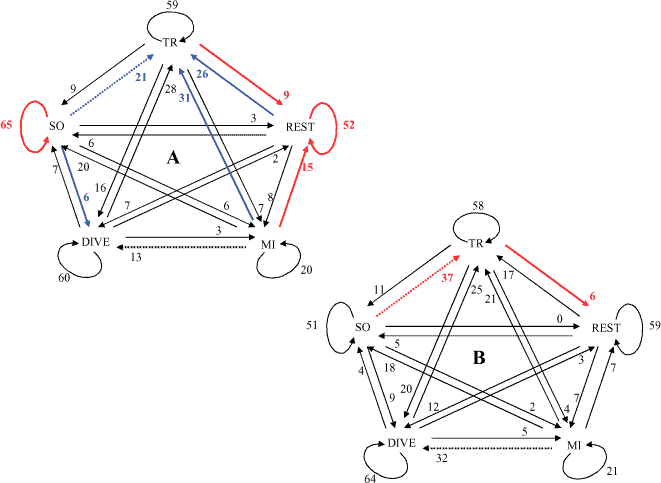
Comparison of transition probabilitiesThe control chains were very similar in both locations (Fig. 3); only two transitions varied significantly. The significance of the effect of boat interactions was not similar in both fjords, because only two transitions were affected in Milford Sound, whereas eight were significantly changed in Doubtful Sound (Fig. 3). This discrepancy could be explained by the marginally small sample size for the impact chain in Milford Sound. Interestingly, the probability that a dolphin would start traveling after socializing was affected differently (Fig. 3). It increased significantly during boat interactions in Doubtful Sound, whereas it decreased significantly in Milford Sound.
|
Fig. 3. Probabilities to shift from one behavioral state to another when there are no boats interacting with the focal school in Doubtful Sound (A) and Milford Sound (B). The behavioral states represented include traveling (TR), socializing (SO), resting (REST), diving (DIVE), and milling (MI). Values are transition probabilities (percentages) of control chains. Colored transitions are significantly affected by boat interactions. Blue transitions are significantly increased, and red transitions are significantly reduced. Dotted lines represent transitions that differ significantly between the two fjords.
|
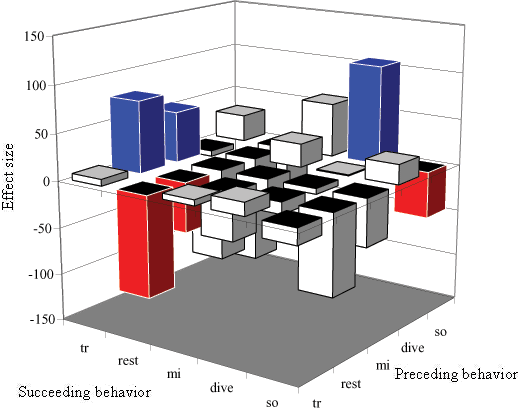
When the effect of location was ignored, as recommended by the log-linear analysis, the effect of boat interactions was not homogeneous over all transitions; some behavioral states were more affected than others. Boat presence changed six transitions significantly (Fig. 4). Three transitions were significantly increased, specifically, those involving traveling and diving as succeeding behavior (Fig. 4). Three transitions were significantly decreased by boat interactions, i.e., those involving either resting or socializing behavior (Fig. 4). The magnitude of the difference in transition probabilities between the two chains is important. The probability of instigating a resting bout from a diving or traveling state was decreased by almost 1.5 times (Fig. 4). The probability of staying in a resting state (prest-rest) decreased by half, as did the probability of continuing to socialize (pso-so). Effect size was based on standardized differences in transition probabilities:
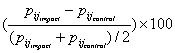 . . | (4) |
|
Fig. 4. The effect of boat presence on transitions in behavioral state in both fjords. The seven transitions with a significant difference (p < 0.05) are colored blue to indicate an increase and red for a decrease. The behavioral states are abbreviated as follows: traveling (tr), resting (rest), milling (mi), diving (dive), and socializing (so).
|
Behavioral budget
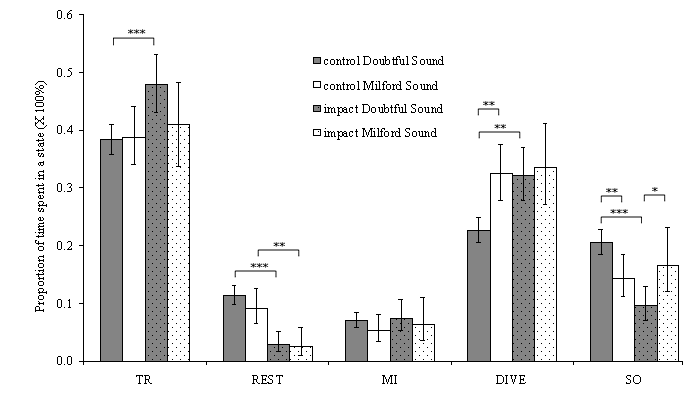
Dolphins spent a similar amount of time socializing regardless of boat presence (Fig. 5) in Milford Sound. In that fjord, only the time spent resting was affected by boat presence (9.1 vs. 2.5%). In Doubtful Sound, all states except milling were significantly affected. Control behavioral budgets differed from one another (Fig. 5). Dolphins in Milford Sound spent more time diving (32.5 vs. 25.5%) and less time socializing (14.3 vs. 21.5%). This is explained by the differences in transition probabilities observed between the two fjords. Dolphins were more likely to go from socializing to traveling and milling to diving in Milford Sound than in Doubtful Sound.
|
Fig. 5. Behavioral budget for Milford Sound (MS) and Doubtful Sound (DS) depending on boat presence. Budgets are given as the proportion of time spent in a given state (sum = 1 across states). Error bars are 95% confidence interval. The significance of the difference between each effect (location and boat impact) is given for each behavioral state above the bars (z-ratio statistic, Fleiss 1981). * = p < 0.05, ** = p < 0.001, *** = p < 0.0001. TR stands for traveling; REST, resting; MI, milling; DIVE, diving; and SO, socializing.
|
Modeling the effects
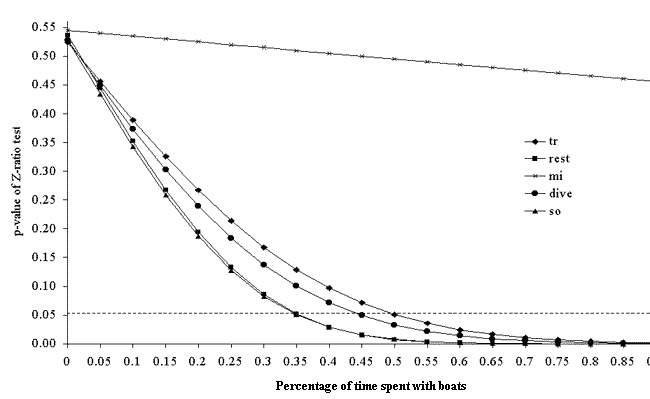
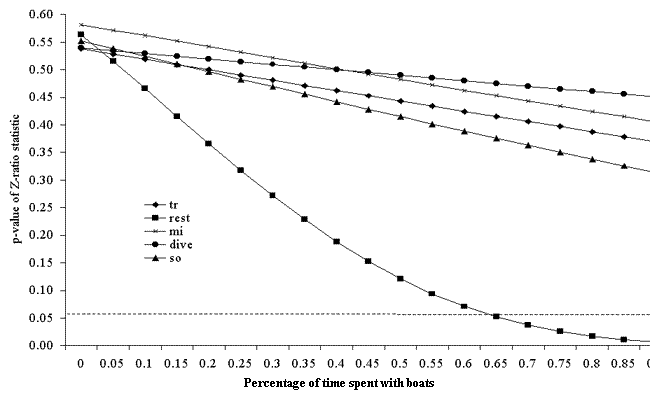
In both fjords, the cumulative behavioral budgets were no different from the control budgets (Figs. 6 and 7). If effects build linearly, in Doubtful Sound dolphins would need to spend 35% of their time with boats for their resting behavior to change significantly (Fig. 6). These dolphins would need to be with boats for 50% of their time for most of their behavior to become significantly different from the control budget (Fig. 6). In Milford Sound, dolphins would need to spend 65% of their time with boats for their cumulative resting behavior to be significantly disturbed (Fig. 7). These estimates assume that the response curve, i.e., the effect of boat impact, stays the same as interaction intensity increases. If the nature of boat-dolphin interactions was similar in both fjords, it would be expected that dolphins in Milford Sound would spend roughly five times more time with boats than do those in Doubtful Sound, because boat traffic in Milford Sound is approximately five times denser. This would therefore mean that dolphins would spend 50–55% of the time they are in Milford Sound with boats. Using the Doubtful Sound model for cumulative behavioral budget (Fig. 6) as an indicator, the cumulative behavioral budget of the Milford Sound population would be significantly affected.
|
Fig. 6. The p-value of the two-tailed Z-test (proportion test) for differences in the percentage of time spent in a given state between the cumulative behavioral budget and the control behavioral budget in Doubtful Sound. The proportion of time spent with boats was modeled to vary from 0 to 100%. The states represented include traveling (tr), resting (rest), milling (mi), diving (dive), and socializing (so).
|
|
Fig. 7. The p-value of the two-tailed Z-test (proportion test) for differences in percentage of time spent in a given state between the cumulative behavioral budget and the control behavioral budget in Milford Sound. The proportion of time spent with boats was modeled to vary from 0 to 100%. The states represented include traveling (tr), resting (rest), milling (mi), diving (dive), and socializing (so).
|
Time needed between two boat interactions
The time elapsed between two boat interactions varied significantly between seasons in Doubtful Sound, ranging from 69.9 to 238.5 min. Moreover, this variation was significantly related to the number of boat trips undertaken each season (F1,5 = 14.371, p = 0.013, R2 = 69.0%). Unexpectedly, the mean period between boat interactions in Milford Sound did not vary with boat traffic (F1,2 = 0.033, p = 0.87) and did not vary seasonally. This period was constant across seasons, averaging 67.6 min (SE = 0.3).
In both fjords, the resting behavior of the dolphins was disrupted and shortened by the presence of boats. In Doubtful Sound, the effect was much stronger, and socializing behavior was also strongly impacted by interactions with boats. The impacts observed in Doubtful Sound indicate a horizontal avoidance of boats, i.e., an increase in traveling behavior during most interactions, but especially when dolphins are socializing or resting. It seems that dolphins in Milford Sound may have taken the additional step of avoiding the area when boat traffic is high (Lusseau et al. 2002, Lusseau 2003c).
This study shows that resting and socializing behavior are sensitive to boat interactions and therefore need to be protected in Doubtful and Milford Sounds. In both places, boat interactions should be minimized during resting bouts. In Doubtful Sound, socializing bouts must be protected as well. Socializing is likely to be directly related to the reproductive output of a population, so that less time for socializing might easily result in lower pregnancy rates. Because the Milford Sound population has been studied for only the past 3 yr, population ecology parameters of this type cannot be reliably compared between the two locations. The proportion of time spent socializing in Milford Sound was much shorter than in Doubtful Sound. This difference may be related to the high boat traffic in Milford Sound. Dolphins may already be altering their behavioral budget while in Milford Sound to cope with the intense boat traffic. The impact on socializing bouts in Milford Sound is not as obvious as in Doubtful Sound, but it is present and warrants the protection of socializing bouts in this fjord as well.
Different strategies for different levels of boat trafficIt is interesting to note that, compared with Doubtful Sound, the impacts of boat interactions in Milford Sound seem to decrease as boat traffic increases. Dolphins in Milford Sound may be avoiding a significantly different cumulative behavioral budget by avoiding Milford Sound at busy times (Lusseau 2003c).
It is also possible that the difference observed between the two fjords is a sign of tolerance in areas in which boat traffic has become more intense. However, this seems unlikely because of the sometimes extreme nature of dolphin-boat interactions in the area, e.g., four individuals in the populations bear the marks of boat collisions and one calf was killed by a boat (Lusseau et al. 2002). Dolphins may be using different avoidance strategies to deal with different levels of boat traffic. Even though boat traffic in Milford Sound is five times higher than in Doubtful Sound, dolphins spend roughly the same amount of time with boats in both fjords.
In Doubtful Sound, the level of boat traffic is relatively low, and dolphins are able to mitigate its impact by moving away, vertically or horizontally, to areas within the fjord with less traffic. High levels of boat traffic on Milford Sound make it difficult for dolphins to avoid encounters either horizontally or vertically. Dolphins in Milford Sound appear to rearrange their behavioral budget around the boat traffic timetable and avoid the area when boat traffic is too intense (Lusseau et al. 2002, Lusseau 2003c). Despite this long-term strategy, some short-term avoidance is still required during resting behavior. This could explain the difference in the control behavioral budgets between the two fjords. Several consequences can be hypothesized from this area avoidance strategy. For example, it could be followed by an increase in predation pressure because dolphins have to spend more time along the coastline. It most likely also affects the energy budget of the population because the dolphins now avoid an area that they used to prefer. If such pressures were present, they could be detected at the population biology level. One would expect a difference in both survival probability and reproductive output, with Doubtful Sound being better off than Milford Sound. It is not currently possible to define reliable estimates of survival and reproductive rates in Milford Sound to compare with those of the better known Doubtful Sound population. However, such information should be collected in the future to allow for such comparisons.
Other studies have emphasized the link between responses to boat presence and predation avoidance strategies (Williams et al. 2002). These strategies have been extensively studied (Howland 1974) and show their relationship to energetic cost/benefit ratios, i.e., making the choice between fight and flight. Switching from a short-term behavioral response to area avoidance has been linked to increasing boat traffic for killer whales (Trites et al. 1995, Williams et al. 2002). This scenario seems to be present in Milford Sound. Because of the density of boats and the behavior these boats display around dolphins (Lusseau et al. 2002), bottlenose dolphins appear to have no choice but to avoid Milford Sound at times when boat traffic is intense. The impact of tourism activities is therefore biologically significant in Milford Sound because tourism forces the population to alter the way in which it uses its home range on a long-term basis. Such an effect has not been observed in Doubtful Sound because the intensity of boating activities is lower than the threshold at which a long-term avoidance strategy is triggered.
When is it too much?It appears that the frequency of dolphin-boat interactions in Doubtful Sound is still strongly related to the presence of boats in the area. From this analysis, it does not appear that dolphins are actively trying to stay away from boats. On the other hand, the same results in Milford Sound reinforce previous findings (Lusseau et al. 2002, Lusseau 2003c), that dolphin-boat interactions are not related to boat traffic. It seems that dolphins actively avoid boat interactions and try to maintain them below a certain level. It appears that dolphins in Milford Sound need at least 68 min between two boat interactions. I therefore hypothesize that this period is the threshold at which it is too energetically costly to engage in short-term displacement. If dolphin-boat interactions occur less than 68 min apart, the area avoidance strategy appears to be the only option that allows the population to keep the time between interactions to more than 68 min. As of the summer of 2002, dolphins in Doubtful Sound spent on average 69 min between two boat interactions. This value is very close to the threshold determined in Milford Sound, and it is a good reason why there should be no further development of the tourism industry in the area. It also means that it is quite probable that dolphins will most likely spend more time in Milford Sound if the frequency of interaction with boats is decreased.
This threshold is linked to the energetic budget of the two populations. Both these populations live at the southern extreme of the species' range and therefore need to use more of their total energy available to live than other populations do. Therefore, it would be difficult to extrapolate these findings to populations living in more temperate climates. Further theoretical and field research is necessary to understand how the threshold observed varies with population ecology and other factors.
Responses to this article are invited. If accepted for publication, your response will be hyperlinked to the article. To submit a comment, follow this link. To read comments already accepted, follow this link.
This research was funded by the New Zealand Department of Conservation, Southland Conservancy. The New Zealand Whale and Dolphin Trust and the Departments of Zoology and Marine Sciences of the University of Otago provided additional financial and technical support. This study could not have been carried out without the help of many volunteers. I would also like to thank the Marine Mammal Research Group of the University of Otago, especially Liz Slooten, Steve Dawson, Oliver J. Boisseau, and Susan Mærsk Lusseau for their continuous support, as well as Hal Caswell for his input on the application of this analytical technique. Constructive comments from two anonymous referees helped improve this manuscript. The home range of the Milford population was reconstructed thanks to information provided by K. David Rundgren (University of Otago), Peter Archibald (Hollyford Track Guided Walks), Murray Gunn, and Marty Taylor. I would like to thank Real Journeys Ltd., especially Paul Norris and Frank Wells, as well as Paul Stewart for their continuous help and support in the field.
Altmann, J. 1974. Observational study of behavior: sampling methods. behavior 49:227-267.
Anderson, D. R., K. P. Burnham, and W. L. Thompson. 2000. Null hypothesis testing: problems, prevalence, and an alternative. Journal of Wildlife Management 64(4):912-923.
Bearzi, G., G. Notarbartolo-di-Sciara, and E. Politi. 1997. Social ecology of bottlenose dolphins in the Kvarneri (Northern Adriatic Sea). Marine Mammal Science 13:650-668.
Bejder, L., S. M. Dawson, and J. A. Harraway. 1999. Responses by Hector's dolphins to boats and swimmers in Porpoise Bay, New Zealand. Marine Mammal Science 15(3):738-750.
Caswell, H. 2001. Matrix population models. Sinauer, New York, New York, USA.
Corkeron, P. J. 1995. Humpback whales (Megaptera novaeangliae) in Hervey Bay, Queensland: behavior and responses to whale-watching vessels. Canadian Journal of Zoology 73:1290-1299.
Fleiss, J. L. 1981. Statistical methods for rates and proportions. Wiley, New York, New York, USA.
Guttorp, P. 1995. Stochastic modeling of scientific data. Chapman and Hall, London, UK.
Hill, M. F., and H. Caswell. 2001. The effects of habitat destruction in finite landscapes: a chain-binomial metapopulation model. Oikos 93:321-331.
Howland, H. C. 1974. Optimal strategies for predator avoidance: the relative importance of speed and manoeuvrability. Journal of Theoretical Biology 47:333-350.
Janik, V. M., and P. M. Thompson. 1996. Changes in surfacing patterns of bottlenose dolphins in response to boat traffic. Marine Mammal Science 12:597-602.
Kruse, S. 1991. The interactions between killer whales and boats in Johnstone Strait, B.C. Pages 149-159 in K. S. Norris and K. Pryor, editors. Dolphin societies: discoveries and puzzles. University of California Press, Berkeley, California, USA.
Lusseau, D. 2002. The state of the scenic cruise industry in Doubtful Sound in relation to a key natural resource: bottlenose dolphins. Pages 106-117 in Proceedings of the Ecotourism, Wilderness and Mountain Tourism Conference (Dunedin, 2002). Centre for Tourism, University of Otago, Dunedin, New Zealand.
Lusseau, D. 2003a. The effects of tour boats on the behavior of bottlenose dolphins: using Markov chains to model anthropogenic impacts. Conservation Biology 17:1785-1793.
Lusseau, D. 2003b. Male and female bottlenose dolphins (Tursiops spp.) have different strategies to avoid interactions with tour boats in Doubtful Sound, New Zealand. Marine Ecology Progress Series 257:267-274.
Lusseau, D. 2003c. The effects of tourism activities on bottlenose dolphins in Fiordland. Dissertation. University of Otago, Dunedin, New Zealand.
Lusseau, D., K. Schneider, O. J. Boisseau, P. Haase, E. Slooten, and S. M. Dawson. 2003. The bottlenose dolphin community of Doubtful Sound features a large proportion of long-lasting associations. Behavioral Ecology and Sociobiology 54:396-405.
Lusseau, D., E. Slooten, S. M. Dawson, and J. Higham. 2002. The effects of tourism activities on bottlenose dolphins (Tursiops spp.) in Fiordland: working towards a sustainable solution; final report to the New Zealand Department of Conservation. New Zealand Department of Conservation, Wellington, New Zealand.
Mann, J. 2000. Unraveling the dynamics of social life. Pages 45-64 in J. Mann, R. C. Connor, P. L. Tyack, and H. Whitehead, editors. Cetacean societies: field studies of dolphins and whales. University of Chicago Press, Chicago, Illinois, USA.
Moberg, G. P. 2000. Biological response to stress: implications for animal welfare. Pages 1-21 in G. P. Moberg and J. A. Mench, editors. The biology of animal stress. CAB International, Wallingford, UK.
Newcombe, R. G. 1998. Two-sided confidence intervals for the single proportion: comparison of seven methods. Statistics in Medicine 17:857-872.
Nowacek, S. M., R. S. Wells, and A. R. Solow. 2001. Short-term effects of boat traffic on bottlenose dolphins, Tursiops truncatus, in Sarasota Bay, Florida. Marine Mammal Science 17(4):673-688.
Shane, S. H. 1990. Behavior and ecology of the bottlenose dolphin at Sanibel Island, Florida. Pages 245-266 in S. Leatherwood and R. R. Reeves, editors. The bottlenose dolphin. Academic Press, New York, New York, USA.
Tanner, J. E., T. P. Hughes, and J. H. Connell. 1994. Species coexistence, keystone species and succession in coral assemblages: a sensitivity analysis. Ecology 75:2204-2219.
Trites, A. W., W. Hochachka, and S. K. Carter. 1995. Killer whales and vessel activity in Robson Bight from 1991 to 1994; report to the Canadian Ministry of Environment, Land and Parks. BC Parks, Strathcona District, Parksville, British Columbia, Canada.
Van Parijs, S. M., and P. J. Corkeron. 2001. Boat traffic affects the acoustic behavior of Pacific humpback dolphins, Sousa chinensis. Journal of the Marine Association U.K. 81:533-538.
Williams, R., A. W. Trites, and D. Bain. 2002. Behavioral responses of killer whales (Orcinus orca) to whale-watching boats: opportunistic observations and experimental approaches. Journal of Zoology 256:255-270.
Address of Correspondent:
David Lusseau
University of Aberdeen
Lighthouse Field Station
George Street,
Cromarty, Ross-shire, UK IV11 8YJ
Phone: (44) 1381-600-548
Fax: (44) 1381-600-548
d.lusseau@abdn.ac.uk
![]()