|
|
|
Copyright © 2000 by The Resilience Alliance
The following is the established format for referencing this article:
McDaniel, C. J., L. B. Crowder, and J. A. Priddy. 2000. Spatial dynamics of sea turtle abundance and shrimping intensity in the U.S. Gulf of Mexico. Conservation Ecology 4(1): 15. [online] URL: http://www.consecol.org/vol4/iss1/art15/
A version of this article in which text, figures, tables, and appendices are separate files may be found by following this link.
Report Spatial Dynamics of Sea Turtle Abundance and Shrimping Intensity in the U.S. Gulf of Mexico Carrie J. McDaniel, Larry B. Crowder, and Jeffery A. Priddy
Duke University Marine Laboratory
- Abstract
- Introduction
- Methods
- Results
- Discussion
- Responses to this Article
- Acknowledgments
- Literature Cited
- Appendix 1
In order to examine the scientific feasibility of area closures for sea turtle protection, we determined the spatial dynamics of sea turtles for the U.S. Gulf of Mexico by analyzing National Marine Fisheries Service (NMFS) aerial survey data in September, October, and November of 1992, 1993, and 1994. Turtle sightings were grouped into depth zones and NMFS fishery statistical zones, and strip transect methods were used to estimate the relative abundance of sea turtles in each subzone. Average shrimping intensity was calculated for each subzone for all months of 1992, 1993, and 1994, as well as for the months and locations of the aerial survey. The spatial overlap of sea turtle abundance and shrimping intensity suggested regions where interactions are likely to occur. Sea turtles were observed at much higher rates along the coast of Florida than in the Western Gulf; the highest density of sea turtles was observed in the Florida Keys region (0.525 turtles/km2). Shrimping intensity was highest in the Western Gulf along the coast of Texas and Louisiana, for both annual and fall estimates. Among alternative management scenarios, area closures in conjunction with continued Turtle Excluder Device (TED) requirements would probably best prevent sea turtles from future extinction. By implementing shrimping closures off of South Padre Island, Texas, a potential second nesting population of Kemp's ridleys (Lepidochelys kempi) could be protected. Closing waters where shrimping intensity is low and sea turtle abundance is high (e.g., South Florida waters) would protect sea turtles without economically impacting a large number of shrimpers.
KEY WORDS: density estimates, endangered species, GIS, Gulf of Mexico, Kemp's ridley, management, sea turtles, shrimp fishing, spatial dynamics, strip transect methods.
Published: April 13, 2000
All species of sea turtles in U.S. waters are listed on the Endangered Species Act of 1973 as endangered or threatened. The five species found in the U.S. Gulf of Mexico are the loggerhead (Caretta caretta), green (Chelonia mydas), hawksbill (Eretmochelys imbricata), Kemp's ridley (Lepidochelys kempi), and leatherback (Dermochelys coriacea) sea turtles. The most common are the loggerhead and Kemp's ridley, but the latter is the most endangered sea turtle worldwide. Kemp's ridley turtles nest almost exclusively at one nesting location at Rancho Nuevo, Mexico, but occasional nests have been found in Veracruz, Mexico, and from Mustang Island, Texas, to Isla Aguada in the Yucatan (Ross et al. 1989).
Bycatch in shrimp trawl fisheries has been determined to be a major source of mortality for large juvenile and adult sea turtles (Henwood and Stuntz 1987, Murphy and Hopkins-Murphy 1989). In 1990, a National Research Council study estimated that 5,000 to 55,000 sea turtles died each year in shrimping activities, more than any other human-induced source of mortality (Magnuson et al. 1990). The influence of shrimp trawling on sea turtle survival is especially noteworthy in the Gulf of Mexico, which it is the site of the largest shrimp fishery in the United States (Magnuson et al. 1990). In the Gulf, most of the shrimp are caught in Louisiana and Texas waters from April to October (Magnuson et al. 1990, Caillouet et al. 1991), and most shrimping activity is concentrated close to shore (Gallaway et al. 1995). Similarly, sea turtles are also found close to shore during the summer when the waters are warm (Hildebrand 1983, Byles 1989, Lohoefener et al. 1990, Manzella and Williams 1992).
An early loggerhead population model suggested that improved survival in the large juvenile and subadult stages would contribute most to population recovery (Crouse et al. 1987). Further, Turtle Excluder Devices (TEDs), which were introduced by National Marine Fisheries Service (NMFS), were developed to provide appropriate protection for these critical stages. Regulations implementing TEDs were passed in 1989, and in December of 1992, TEDs were required year-round in offshore waters of the United States. Since 1995, TED compliance has been high (about 97.6%) in the Gulf of Mexico, and most shrimpers are properly installing and using TEDs (M. Johnson, U.S. Coast Guard, personal communication). Yet, sea turtle mortality remains a problem. Significantly increased strandings (dead turtles washed up on the beach), including a high proportion of Kemp's ridleys, occurred throughout the Gulf in 1994, and strandings were again high in 1995, especially in Louisiana and Texas. Strandings previously have been correlated with shrimping activity (Magnuson et al. 1990, Caillouet et al. 1991) and perhaps with changes in population size (Crowder et al. 1995).
A preliminary Kemp's ridley population model (Turtle Expert Working Group 1998) suggests a post-1990 improvement in survival of large juveniles and adult Kemp's ridleys, but to date, no analyses of empirical data have demonstrated a TED effect in reducing mortality or strandings in Kemp's ridleys (Caillouet et al. 1996). Increased strandings of Kemp's ridleys in South Texas have been a concern to those attempting to establish a second nesting population at Padre Island National Seashore (Shaver and Caillouet 1998). Some have suggested that TEDs alone may not provide adequate protection for sea turtles in the Gulf of Mexico, and that managers should evaluate the potential for seasonal or spatial closures as a supplement (or alternative) to TEDs (Gallaway et al. 1995, Shaver and Caillouet 1998).
Time and area closures have been effective in solving conservation problems, particularly for nonmigratory species, in terrestrial systems. Clearly, evaluating the potential for such closures is more complicated for migratory species or marine organisms that spend a large portion of their lives under water. Sea turtles can migrate over vast distances, but the spatial distribution and seasonal movements of sea turtles are poorly understood. The shrimp trawl fleet, a major threat to sea turtle survival, also moves about seasonally (Caillouet et al. 1991). The Turtle Expert Working Group (1998) realized the problems associated with the spatial and temporal dynamics of sea turtles and shrimpers, and called for better in-water surveys to understand both population trends and spatial interactions.
One technique used to estimate the abundance of sea turtles in the water is an aerial survey. Aerial surveys may be the only practical approach to determining the relative distribution and abundance of certain animals over extensive areas (Bayliss 1986). Several research studies have used aerial survey techniques successfully to estimate sea turtle abundance and location (Shoop and Kenney 1992, Musick et al. 1994, Epperly et al. 1995). At-sea aerial observations cannot be assumed to provide complete population estimates for marine turtles, but they are useful for mapping the relative abundance and location of sea turtles.
In this project, the primary goal was to determine the relative spatial pattern of sea turtle abundance and shrimping intensity throughout the Gulf of Mexico, and to outline possibilities for future research and management. We were also interested in examining the distribution of surfacing sea turtles over water of different depths, because limited satellite telemetry of individuals suggests that more sea turtles inhabit nearshore waters (Hildebrand 1983, Byles 1989, Lohoefener et al. 1990, Gallaway et al. 1995, Renaud 1995). In this paper, we compared fall sea turtle distributions in the Gulf of Mexico to the spatial distribution of shrimping activity during fall and over the entire year.
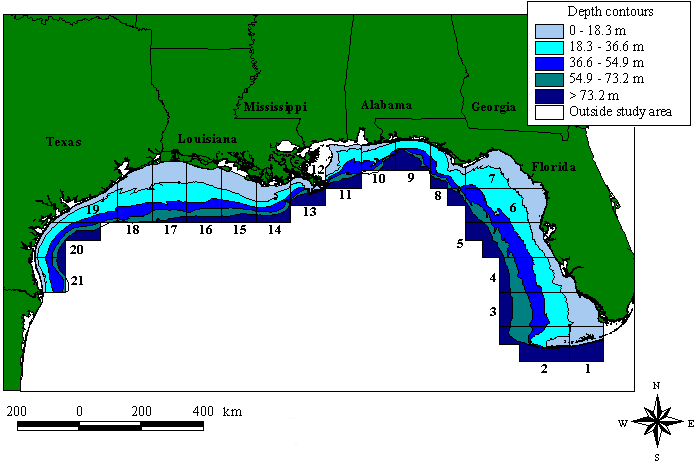
The study area was the U.S. Gulf of Mexico, extending approximately 80o to 98o W longitude and 24o to 31o N latitude. The U.S. Gulf is divided into 21 statistical zones (Fig. 1), determined by NMFS to simplify reporting of fisheries landings and effort. This study used depth contours in 18.3-m (10-fathom) intervals and each "bin" created by the intersection between the statistical zones and depth contours represented a subzone. For each subzone, sea turtle densities and shrimping intensity were estimated.
|
Fig. 1. The U.S. Gulf of Mexico study area and depth contours. Boxes represent NMFS statistical zones and their corresponding numbers. Depth contours are in 18.3-m (10-fathom) intervals.
|
Sea turtle sightings
Sea turtle observations were obtained via aerial surveys conducted by NMFS throughout the U.S. Gulf of Mexico. From 1992 to 1994, GOMEX (Gulf of Mexico) surveys were flown to estimate cetacean abundance and distribution in the Gulf coastal and continental shelf waters. During each survey, sea turtle sightings were also recorded. GOMEX92 was conducted from 13 September to 24 October 1992, from Brownsville, Texas to Lafayette, Louisiana. GOMEX93 was conducted from 17 September to 19 October 1993, from Lafayette, Louisiana to Cedar Key, Florida, and GOMEX94 was performed from 28 September to 9 November 1994, from Cedar Key, Florida to Key West, Florida (Anonymous 1992). The transects extended from the shoreline to approximately 9.3 km past the 193-m (100-fathom) isobath. Although we would have preferred to have data on sea turtle and shrimp fleet distributions across the peak of the shrimping season (April to October; Magnuson et. al 1990, Gallaway et al. 1995), these are the only comprehensive aerial survey sea turtle data available.
The survey platform was a NOAA-owned DeHavilland DH-6 Twin Otter aircraft with a plastic bubble window on each side of the plane. The location and observations were compiled in a portable computer interfaced with a global positioning system (GPS). All GOMEX surveys were conducted at an altitude of 229 m and an approximate speed of 220 km/h (Blaylock 1993, Blaylock and Hoggard 1994).
Because all sea turtle species are protected in U.S. waters and there were comparatively few total sightings, we did not differentiate among sea turtle species in the aerial surveys. This avoided any misidentification related to platform altitude or speed. Aerial survey transects were divided into smaller transects, based on the depth contours. The sea turtle sightings were grouped into 18.3-m depth bins, and sightings beyond 73.2 m were combined (e.g., 0-18.3; 18.3-36.6; 36.6-54.9; 54.9-73.2; >73.2 m).
Strip transect methodologySea turtle abundance for each subzone was calculated using strip-transect methodology. The basic premise behind this method is that a strip of a specific width is defined in the aerial survey process, and only the animals within this strip are counted. The strip width was determined from a histogram of all the perpendicular sighting distances. The perpendicular sighting distance is defined as the distance from the flight line to the observed organism (Burnham et al. 1980). In this study, the effective strip width was 50-250 m on both sides of the survey platform. By choosing a starting strip distance of 50 m from the survey platform, we reduced the probability that a sea turtle was missed because of proximity to the plane. Close proximity to the plane could result in undercounting sea turtles from the acute observer viewing angle or from turtles diving to avoid the noisy aircraft (Epperly et al. 1995). The ending distance of 250 m was determined by the decline in perpendicular sighting distance values. This strip width reduces the probability of missing a sea turtle due to the distance from the plane (reduced detection). Only sightings within this strip width were used for the relative density estimates. Appendix 1 details the procedure used to calculate density estimates.
Shrimping intensityGulf of Mexico shrimping effort was collected through a port agent program, run by NMFS. Two types of data were used to estimate shrimping effort: dealer data (landings through a recognized dealer) and interview data by NMFS port agents (actual interviews with captains following a fishing trip; Nance 1993). The shrimping effort data that we received from NMFS Galveston were measured in number of days fished (24 hours of actual fishing), and the shrimping effort value for each subzone was standardized into number of days fished per square kilometer. The area of each subzone was previously determined by Patella (1975).
Offshore shrimping intensities were estimated for each 18.3-m interval of depth from 0 m to the end of the statistical zone. Shrimping intensity in waters deeper than 73.2 m was combined. Statistical zone 12 was excluded from this analysis because all waters in zone 12 are inside a string of barrier islands (Chandeleur Islands), and are therefore considered to be inshore. Therefore, this project analyzed 100 subzones in the Gulf (5 depth bins x 20 statistical zones = 100 total subzones).
Two analyses were performed; one included all of the shrimping months (January through December 1992 to 1994) and the second considered the averaged shrimping intensity only for the months and locations of the aerial surveys (zones 1-6 for September to November 1994; zones 7-15 for September to October 1993; and zones 16-21 for September to October 1992). It is possible that the fall distribution and relative abundance of sea turtles are affected by shrimping effort in the preceding seasons as well as the fall.
Spatial analysisThe sea turtle abundance and shrimping intensity were entered into a Geographical Information System (GIS) to describe the spatial patterns throughout the Gulf. The GIS packages used were UNIX-based ArcInfo and ArcView. The original depth contours, statistical zones, and detailed Gulf of Mexico coastline files were obtained from NMFS Galveston.
All cumulative estimates of sea turtle abundance and shrimping intensity were divided into thirds to give low, medium, and high categories. Each value was considered in the ranking (e.g., all "0" values), resulting in nine different combinations of shrimping intensity and sea turtle abundance. This ranking system should allow for qualitative comparisons among high and low areas of shrimping, as well as high and low sea turtle abundance.
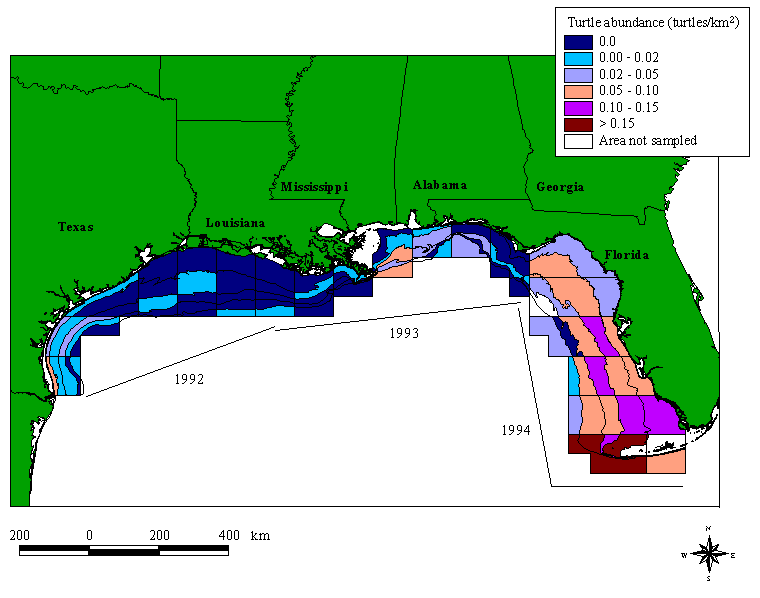
The relative abundance of sea turtles sighted in the GOMEX surveys varies with fishery statistical zone and depth throughout the Gulf (Fig. 2). In general, the relative abundance of turtles sighted is higher in the Eastern Gulf than in the Western Gulf. The southern Florida zones (zones 1-3) apparently have a higher relative abundance of sea turtles than any other region. The west coast of Florida has sea turtle densities that are, on average, 60 times higher than in the Western Gulf, but three times lower than in the southern Florida areas. Overall densities remain high (0.047 turtles/km2) through the Big Bend region of Florida (zone 7), but abundance is reduced off Mississippi and Alabama. West of the Mississippi River, observed sea turtle abundance is extremely low, with no turtles sighted in many of the subzones. In southern Texas (zones 20 and 21), however, average sea turtle abundance is about 20 times higher than in the other Western Gulf zones.
|
Fig. 2. Sea turtle abundance for September, October, and November 1992, 1993, and 1994 in the U.S. Gulf of Mexico. Abundances were based on aerial survey sightings; dated lines represent the year and location of the aerial survey flight. Depth contours are in 18.3-m (10-fathom) intervals and the boxes are NMFS statistical zones. The intersection of the statistical zone and depth contour represents a subzone.
|
Shrimping intensity
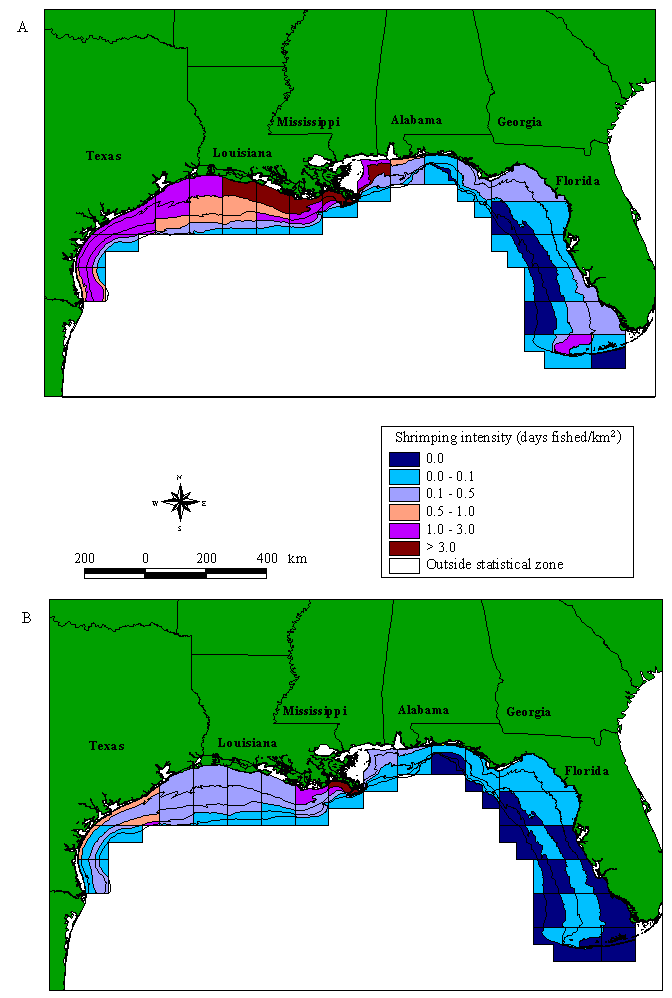
Gulf shrimping intensity over the whole season shows a very distinct pattern (Fig. 3A). The Western Gulf has an extremely high level of shrimping, as does zone 11 on the eastern side of the Mississippi River. In almost all of the western subzones, shrimping does not fall below 0.439 days fished/km2. To the east of zone 11, shrimping intensity decreases; the Florida shrimp fishery appears to be negligible in comparison to the Western Gulf. One area of high shrimping intensity does occur outside the Dry Tortugas Shrimp Sanctuary in southern Florida (zone 2 from 18.3 to 36.6 m). Throughout the Gulf, shrimp fishing is generally found close to shore, in waters < 54.9 m. This general assessment of shrimping intensity is consistent with the landings data that derive primarily from Louisiana and Texas waters (Caillouet et al. 1991).
|
Fig. 3. Shrimping intensity in the U.S. Gulf of Mexico. (A) Average all-year shrimping intensity for 1992, 1993, and 1994. (B) Average shrimping intensity for zones 1-6 for September to November 1994; zones 7-15 for September to October 1993; and zones 16-21 for September to October 1992. Depth contours, NMFS statistical zones, and subzones are defined as in Fig. 2.
|
Shrimping intensity in the fall is somewhat lower than the all-year values, but the spatial distribution of shrimping effort appears to be similar (Fig. 3B). Shrimping remains moderately high throughout the western Gulf, with intensive shrimping off the Texas coast (zones 18-20) and east Louisiana (zone 13-14). The Eastern Gulf has a noticeably reduced shrimping intensity.
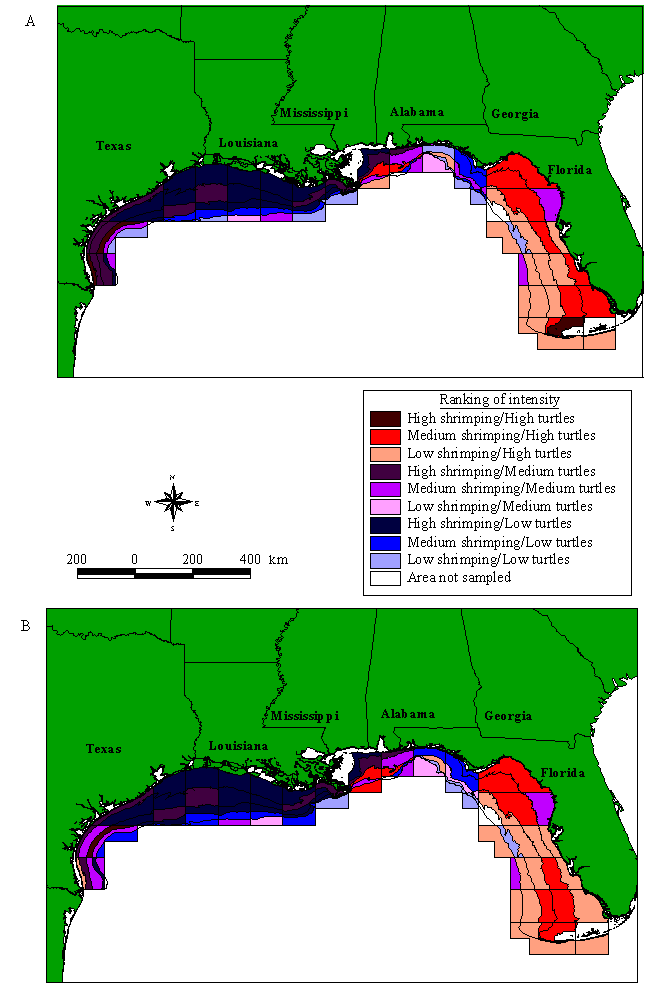
Sea turtle relative abundance and shrimping intensityRankings of sea turtle relative abundance and all-year shrimping intensity are shown in Fig. 4A. The southern Florida region of the Gulf has relatively high sea turtle abundance and low shrimping intensity. One exception is the area around the Dry Tortugas Shrimp Sanctuary (zones 1 and 2 from 18.3 to 36.6 m); these subzones have both high shrimping and high sea turtle abundance. In most of the Western Gulf (zones 13-19), shrimping intensity is consistently high, whereas relative sea turtle abundance is uniformly low. However, in the southern Texas region (zones 20 and 21), the relative abundance of sea turtles increases and shrimping intensity is high (Fig. 5A).
|
Fig. 4. Shrimping intensity and sea turtle abundance in the U.S. Gulf of Mexico. (A) Shrimping intensity averaged over all months in 1992, 1993 and 1994. (B) Shrimping intensity averaged for zones 1-6 for September to November 1994; zones 7-15 for September to October 1993; and zones 16-21 for September to October 1992. Depth contours, NMFS statistical zones, and subzones are defined as in Fig. 2.
|
|
Fig. 5. Shrimping intensity and sea turtle abundance in the southern Texas Gulf of Mexico waters (zones 20 and 21). (A) Shrimping intensity averaged over all months in 1992, 1993, and 1994. (B) Shrimping intensity averaged for zones 1-6 for September to November 1994; zones 7-15 for September to October 1993; and zones 16-21 for September to October 1992. Depth contours, NMFS statistical zones, and subzones are defined as in Fig. 2.
|
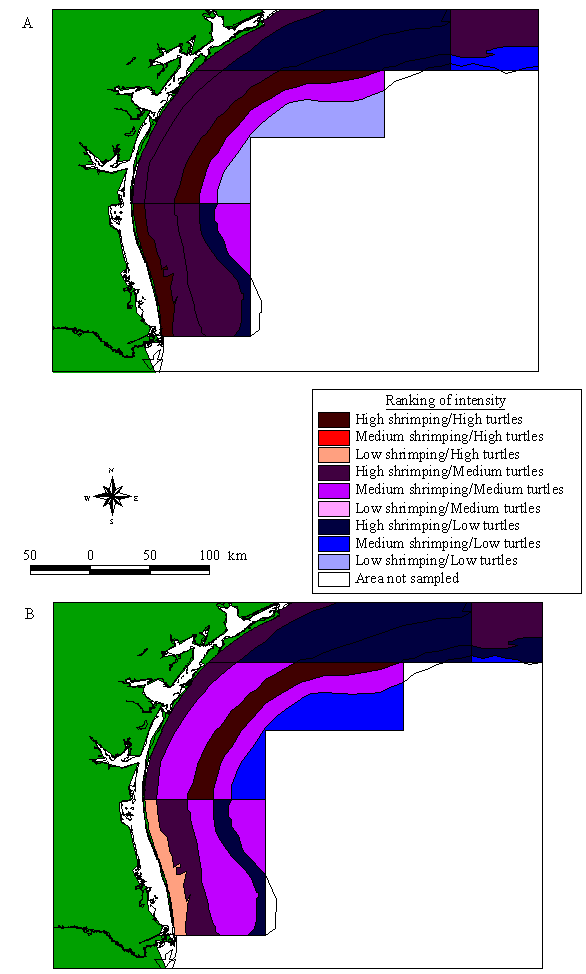
By plotting sea turtle densities and all-year shrimping intensity for each statistical zone, an apparent correlation can be depicted (Fig. 6). Shrimping intensity is low in the Eastern Gulf and then rapidly increases west of the Mississippi River delta (zone 13). Intensities in the other Western Gulf zones are not as high as in zone 13, but they are much higher than in the Eastern Gulf. In contrast, the relative abundance of sea turtles is highest in zones with lower shrimping effort. Turtle abundance is high in southern Florida (zones 1 and 2), and then drops steadily down to zone 8. After the slight jump in abundance in zone 11, relative sea turtle densities are reduced in the remainder of the Western Gulf, except for higher turtle sightings in zones 20 and 21.
|
Fig. 6. Shrimping intensity and sea turtle abundance divided into NMFS statistical zones in the U.S. Gulf of Mexico. Shrimping intensity in 1992, 1993, and 1994 is shown both averaged over the fall months (diamonds) and averaged over all months (triangles).
|
If we compare the relative distribution of turtles to shrimping intensity only in the fall, the general patterns persist (Figs. 4B and 5B). The percentage of total fall shrimping intensity by statistical zone is similar to the annual pattern (Fig. 6).
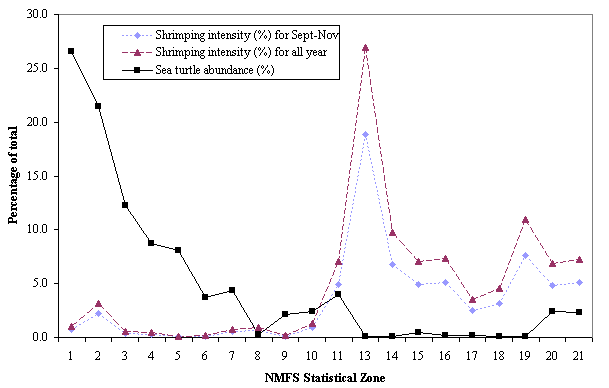
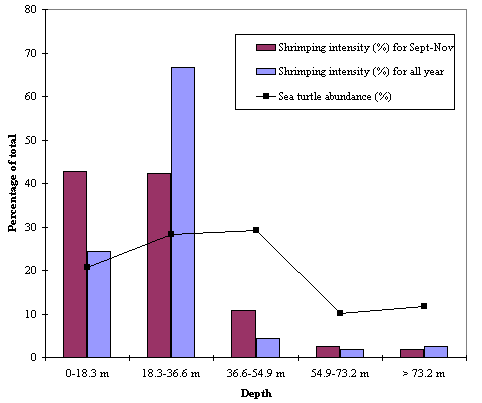
Sea turtle abundance and shrimping intensity by depth zonesOne goal of our study was to examine the relative abundance of sea turtles and shrimping intensity by depth contours. We considered the Eastern and Western Gulf zones separately because relative abundance of turtles was much higher in the Eastern Gulf and could have skewed the Western Gulf distribution. All-year shrimping intensity in the Eastern Gulf appears to be higher in the 18.3-36.6 m category than in any other depth zones (Fig. 7). Nearshore (0-18.3 m) sea turtle abundance in the Eastern Gulf is slightly lower than in the mid-depths, but at depths > 54.9 m, sea turtle relative abundances seem to decline. Western Gulf shrimping intensities tend to be highest nearshore and gradually taper off with depth. Nearshore sea turtle abundances are proportionately higher than in offshore Western Gulf subzones, with the greatest density of sea turtles found in the 0-18.3 m zone (Fig. 8).
|
Fig. 7. Eastern Gulf shrimping intensity and sea turtle abundance divided into depth bins in the U.S. Gulf of Mexico. Shrimping intensity in 1992, 1993 and 1994 is shown both averaged over all months and averaged over the fall months. Depth bins are in 18.3-m (10-fathom) intervals.
|
|
Fig. 8. Western Gulf shrimping intensity and sea turtle abundance divided into depth bins in the U.S. Gulf of Mexico. Shrimping intensity in 1992, 1993 and 1994 is shown both averaged over all months and averaged over the fall months. Depth bins are in 18.3-m (10-fathom) intervals.
|
Comparing the spatial distribution of shrimping intensity to the distribution of sea turtles provides insight into potential areas of intensive interaction. However, it is not known with certainty why sea turtles and shrimp fishing intensity are distributed this way. Sea turtle distribution is most probably a result of several factors. Sea turtles could be congregating along the Florida coast because the habitat is more suitable. Sea turtles forage in a variety of habitats, but primary foraging areas are often located on relatively shallow shelves and in warm temperatures (e.g., 25-28o C for loggerheads and Kemp's ridleys; Spotila et al. 1997). The coast of Florida has a wider continental shelf, more seagrasses, and shallower water than the Texas coast; hence, its foraging habitat quality could be higher.
Another possible contributor to the observed spatial distributions is the periods of summer hypoxia in bottom waters off Louisiana (Renaud 1985). Oxygen-deficient conditions do not directly threaten air-breathing sea turtles, but they do affect the distribution of benthic shrimp and other crustaceans that are primary prey for Kemp's ridleys and loggerheads (Gallaway et al. 1995). A hypoxic area would contain a limited biological community and reduced prey; without an adequate food supply, sea turtles would be unlikely to inhabit these locations. Further, if shrimp and sea turtles accumulate at the edge of such a feature (e.g., inshore of the hypoxic zone), interactions with nearshore shrimping activities could be intensified (Craig et al. 2000).
One important result of our study involves the distribution of sea turtles and shrimping off the South Texas coast. Recently, managers have made a strong case for closing the waters off Padre Island National Seashore, Texas, at least during the turtle nesting season (Shaver and Caillouet 1998). The highly endangered Kemp's ridley turtle has nested primarily on one beach in Mexico (Rancho Nuevo), but, partly because of previous headstarting efforts (Magnuson et al. 1990, Woody 1990, Frazer 1992, Heppell et al. 1996), several Kemp's ridleys have nested on Padre Island over the last three years. This beach has the potential to become a second nesting site, but many large juvenile and adult female Kemp's ridleys continue to strand on Texas beaches. Regardless of whether the increase in nesting females at Padre Island is due to headstarting or simply to population growth, developing a second nesting site at Padre Island would help to insure Kemp's ridleys against extinction. If Kemp's ridleys continue to nest exclusively in one geographic location, the species remains at risk due to a local catastrophe (e.g., a hurricane). Our study shows that both shrimping intensity and sea turtle abundance are relatively high in South Texas, at least in the fall (Fig. 5). If these patterns are consistent throughout the shrimping season, a seasonal closure of these waters to shrimping could contribute strongly to the development of a second nesting colony for Kemp's ridleys.
We also clearly identified a relatively high abundance of sea turtles and low shrimping intensity in South Florida waters. Although we did not distinguish species in our analysis, these southern Florida zones also contain a relatively high diversity of sea turtle species. In addition to loggerheads, Kemp's ridleys, and leatherbacks, which were identified in the original GOMEX aerial survey data throughout the Gulf, South Florida also includes substantial numbers of green and hawksbill turtles. Restricting shrimping effort in these areas would protect the high turtle abundance in southern Florida. Furthermore, if managers want to consider seasonal or area closures, closing an area with little shrimping activity and high sea turtle abundance may be easier to implement than in most other Gulf areas.
These recommendations seem relatively straightforward, given our results, but several caveats must be acknowledged. First, we did not attempt to estimate the total population or absolute abundance of sea turtles. In this study, we focused on the relative spatial pattern of abundance throughout the Gulf to point out the zones of potentially intense interactions between shrimping and sea turtles. The second caveat is that the GOMEX aerial surveys were not conducted over the entire shrimping season. The surveys were conducted in September, October, and, in 1994, November, whereas the months of highest shrimping activity occur from June to October (Magnuson et al. 1990). Although the survey period overlaps with a period of active shrimping, many interactions between sea turtles and shrimp trawlers probably occur earlier in the season (Caillouet et al. 1991). Thirdly, the aerial surveys in different years (1992-1994) were conducted in different regions of the Gulf. This could confuse slight regional differences in sea turtle abundance from year to year with real geographic differences; we cannot resolve this with the existing data. However, we compared shrimping intensity in each year only to regions surveyed for turtles in that year. In addition, our analysis provides an overlap of shrimping intensity and sea turtle abundance for September-October only. Several studies have assessed the distribution and movements of sea turtles in the Gulf of Mexico (Rossman et al. 1987, Byles 1989, Ross et al. 1989). By September and October, most of the resident subadults and adults are probably back in their feeding habitats. There seems to be little movement in the fall, at least by turtles large enough to be detected in an aerial survey. Finally, the aerial survey data suggest that the Eastern Gulf has a relatively higher proportion of sea turtles, but relative densities of turtles could be underestimated in the Western Gulf, where turbid waters allow observers to count only turtles at the surface; in the Eastern Gulf, submerged turtles might also have been counted. Other factors including weather, time of day, observer variations, habitat, behavior of the animal, and size of the animal also influence the probability of detection (Buckland et al. 1993), but the frequency of less than optimal surveying conditions should be relatively random from year to year. For example, the probability of a cloudy, choppy day was similar across years.
To verify the regions of potential sea turtle and shrimping interactions, NMFS and other researchers should expand aerial surveys and other in-water assessments of abundance and distribution of sea turtles. The Turtle Expert Working Group (1998) strongly recommended in-water surveys of index locations, particularly as a hedge against depending exclusively upon nesting numbers as the primary index of population trends. Future aerial surveys should be flown during periods of high shrimping activity to determine if sea turtles alter their distributions based on shrimping intensity or season. To provide insight into how sea turtles respond to intense fishing pressure, the survey could be conducted over three different time periods in the same location. For example, a survey in Texas (zone 19) could be conducted before the Texas Closure (late April/early May), during the Texas Closure (June), and after the Texas Closure (late July/early August). This survey would provide a better estimate of how sea turtles respond to high levels of shrimping.
Interactions of protected marine species and fisheries often lead to bycatch and unintended mortality. Turtle Excluder Devices have been required by federal and state managers to reduce sea turtle mortality due to trawling, but despite reportedly good compliance with these regulations, sea turtle strandings in the Gulf have recently been high. Many scientists and managers have begun to consider management alternatives that might supplement the current regulations because current sea turtle protection may not be sufficient under the Endangered Species Act. One management alternative includes seasonal or area closures. Our analysis suggests some options for reducing harmful sea turtle-shrimp trawl interactions, including the protection of a potential second nesting colony for the highly endangered Kemp's ridley sea turtle on Padre Island, Texas, and the protection of high sea turtle abundance off the southern Florida coast.
Responses to this article are invited. If accepted for publication, your response will be hyperlinked to the article. To submit a comment, follow this link. To read comments already accepted, follow this link.
This research was supported by the National Oceanic and Atmospheric Administration, the National Marine Fisheries Service, and the University of North Carolina Sea Grant (R/MER-21 and NOAA/NMFS NA 90AA-D-56847). The aerial survey data were provided by NMFS Pascagoula, and shrimping effort data were obtained from NMFS Galveston. We thank the reviewers, S. Heppell and S. Epperly, for their many insightful comments and suggestions. Recognition also goes to Sheryan Epperly for helping with density estimates, and to Drs. Keith Mullin and Karen Mitchell for assisting with the aerial survey data set interpretation. Benny Gallaway provided valuable insight into the Gulf shrimping industry.
Anonymous. 1992. GOMEX92, Gulf of Mexico regional aerial survey design, September-October 1992. NMFS (National Marine Fisheries Service)-SEFSC (Southeast Fisheries Science Center) Contribution MIA-91/92-72, September 1992.[MIA = Miami, Florida, USA.]
Bayliss, P. 1986. Factors affecting aerial surveys of marine fauna, and their relationship to a census of dugongs in the coastal waters of the Northern Territory. Australian Wildlife Resources 13:27-37.
Blaylock, R. A. 1993. GOMEX92: Survey data documentation report, Gulf of Mexico regional aerial survey, September-October 1992. SEFSC Contribution MIA-92/93-24, January 1993.
Blaylock, R. A., and W. Hoggard. 1994. Preliminary estimates of bottlenose dolphin abundance in southern U.S. Atlantic and Gulf of Mexico continental shelf waters. NOAA Technical Memorandum National Marine Fisheries Service-SEFSC-356.
Buckland, S. T., D. R. Anderson, K. P. Burnham, and J. L. Laake. 1993. Distance sampling: estimating abundance of biological populations. Chapman and Hall, London, UK.
Burnham, K. P., D. R. Anderson, and J. L. Laake. 1980. Estimation of density from line transect sampling of biological populations. Wildlife Monographs 72:1-202.
Byles, R. A. 1989. Satellite telemetry of Kemp's ridley sea turtle, Lepidochelys kempi, in the Gulf of Mexico. Pages 25-26 in S. A. Eckert, K. L. Eckert, and T. H. Richardson, compilers. Proceedings of the Ninth Annual Workshop on Sea Turtle Conservation and Biology. NOAA Technical Memorandum National Marine Fisheries Service-SEFC-232.
Caillouet, C. W. Jr., M. J. Duronslet, A. M. Landry Jr., D. B. Revera, D. J. Shaver, K. M. Stanley, R. W. Heinly, and E. K. Stabenau. 1991. Sea turtle strandings and shrimp fishing effort in the northwestern Gulf of Mexico, 1986-89. U.S. Fishery Bulletin 89:712-718.
Caillouet, C. W., Jr., D. J. Shaver, W. G. Teas, J. M. Nance, D. B. Revera, and A. C. Cannon. 1996. Relationship between sea turtle stranding rates and shrimp fishing intensities in the northwestern Gulf of Mexico: 1986-89 versus 1990-1993. U.S. Fishery Bulletin 94:237-249.
Cochran, W. G. 1977. Sampling techniques. John Wiley, New York, New York, USA.
Craig, J. K., L. B. Crowder, C. D. Gray, C. J. McDaniel, T. A. Henwood, and J. G. Hanifen. 2000. Ecological effects of hypoxia on fish, sea turtles and marine mammals in the Northwestern Gulf of Mexico. In N. N. Rabalais and R.E. Turner, editors. Effects of hypoxia on living resources, with emphasis on the Northern Gulf of Mexico. AGU (American Geophysical Union) Coastal and Estuarine Studies Series.
Crouse, D. T., L. B. Crowder, and H. Caswell. 1987. A stage-based population model for loggerhead sea turtles and implications for conservation. Ecology 68:1412-1423.
Crowder, L. B., S. R. Hopkins-Murphy, and J. A. Royle. 1995. Effects of Turtle Excluder Devices (TEDs) on loggerhead sea turtle strandings with implications for conservation. Copeia 1994(4):773-779.
Epperly, S. P., J. Braun, and A. J. Chester. 1995. Aerial surveys for sea turtles in North Carolina inshore waters. Fishery Bulletin 93:254-261.
Frazer, N. B. 1992. Sea turtle conservation and halfway technology. Conservation Biology 6:179-184.
Gallaway, B. J., J. D. Bryan, L. R. Martin, and J. G. Cole. 1995. Sea turtle and shrimp fishery interactions - is a new management strategy needed? LGL Ecological Research Associates, Bryan, Texas, USA.
Henwood, T. A., and W. E. Stuntz. 1987. Analysis of sea turtle captures and mortalities during commercial shrimp trawling. Fishery Bulletin 85:813-817.
Heppell, S. S., L. B. Crowder, and D. T. Crouse. 1996. Models to evaluate headstarting as a management tool for long-lived turtles. Ecological Applications 6:556-565.
Hildebrand, H. H. 1983. Random notes on sea turtles in the western Gulf of Mexico. Pages 34-40 in D. W. Owens, D. Crowell, G. Dienderg, M. Grassman, S. McCain, Y. Morris, N. Schwantes, and T. Wibbels, editors. Western Gulf of Mexico Sea Turtle Workshop Proceedings. Texas A&M University Publication TAMU-Sg-84-105.
Lohoefener, R., W. Hoggard, K. Mullin, C. Roden, and C. Rogers. 1990. Association of sea turtles with petroleum platforms in the north-central Gulf of Mexico. OCS (Outer Continental Shelf) Study/MMS 90-0025. U.S. Dept. of the Interior, Minerals Management Service, Gulf of Mexico OCS Regional Office, New Orleans, Louisiana, USA.
Magnuson, J. J., J. A. Bjorndal, W. D. DuPaul, G. L. Graham, D. W. Owens, C. H. Peterson, P. C. H. Pritchard, J. I. Richardson, G. E. Saul, and C. W. West. 1990. Decline of sea turtles: causes and prevention. National Research Council, National Academy Press, Washington, D.C., USA.
Manzella, S. A., and J. A. Williams. 1992. The distribution of Kemp's ridley sea turtles (Lepidochelys kempi) along the Texas coast: an atlas. NOAA (National Oceanic and Atmospheric Administration) Technical Report, National Marine Fisheries Service 110, U.S. Department of Commerce, Seattle, Washington, USA.
Murphy, T. M., and S. R. Hopkins-Murphy. 1989. Sea turtle and shrimp fishing interactions: a summary and critique of relevant information. Center for Marine Conservation, Washington, D.C., USA.
Musick, J. A., D. E. Barnard, and J. A. Keinath. 1994. Aerial estimates of seasonal distribution and abundance of sea turtles near the Cape Hatteras faunal barrier. Pages 121-123 in B. A. Schroeder and B. E. Witherington, editors. Proceedings of the Thirteenth Annual Symposium on Sea Turtle Biology and Conservation. NOAA Technical Memorandum, National Marine Fisheries Service-SEFSC-341.
Nance, J. M. 1993. Effort trends for the Gulf of Mexico shrimp fishery. NOAA Technical Memorandum, National Marine Fisheries Service-SEFSC-337.
Patella, F. 1975. Water surface area within statistical subareas used in reporting Gulf coast shrimp data. Marine Fisheries Review 37(12):22-24.
Renaud, M. 1985. Annotated bibliography on hypoxia and its effects on marine life, with emphasis on the Gulf of Mexico. NOAA Technical Report, National Marine Fisheries Service 21, U.S. Department of Commerce, Seattle, Washington, USA.
Renaud, M. 1995. Movements and submergence patterns of Kemp's ridley turtles (Lepidochelys kempi). Journal of Herpetology 29(3):370-374.
Ross, J. P., S. Beavers, D. Mundell, and M. Airth-Kindree. 1989. The status of Kemp's ridley. Center for Marine Conservation, Washington, D.C., USA.
Rossman, I., G. S. Boland, L. Martin, and C. Chandler. 1987. Underwater sightings of sea turtles in the Northern Gulf of Mexico. LGL Ecological Research Associates, Bryan, Texas, USA.
Shaver, D. J., and C. W. Caillouet, Jr. 1998. More Kemp's ridley turtles return to south Texas to nest. Marine Turtle Newsletter 82:1-5.
Shoop, C. R., and R. D. Kenney. 1992. Seasonal distributions and abundances of loggerhead and leatherback sea turtles in waters of the northeastern United States. Herpetological Monographs 6:43-67.
Spotila, J. R., M. P. O'Connor, and F. V. Paladino. 1997. Thermal biology. Pages 297-314 in P. L. Lutz and J. A. Musick, editors. The biology of sea turtles. CRC Press, Boca Raton, Florida, USA.
Turtle Expert Working Group. 1998. An assessment of the Kemp's ridley (Lepidochelys kempi) and loggerhead (Caretta caretta) sea turtle populations in the Western North Atlantic. NOAA Technical Memorandum, National Marine Fisheries Service-SEFSC-409.
Woody, J. B. 1990. Guest editorial: Is "headstarting" a reasonable conservation measure? "On the surface, yes: in reality, no." Marine Turtle Newsletter 50:8-11.
The strip transect methodology used to calculated sea turtle abundances in the U.S. Gulf of Mexico.
Sea turtle observations were converted to densities (with subscript R identifying the base unit for which a density is sought), DR, by dividing the number of sea turtles observed in the subzone (YR) by the total area surveyed in the subzone (MR):
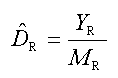 (1)
(1)
 (2)
(2)
where yi is the number of turtles in the ith transect; mi is the area surveyed in the ith transect (km2); and n is the number of strip transects sampled (Cochran 1977). These density estimates were assumed to apply to the whole subzone (depth contour intersected with fishery statistical zone).