






The following is the established format for referencing this article:
Fill, C. T., C. R. Allen, D. Twidwell, and J. F. Benson. 2022. Spatial distribution of bat activity in agricultural fields: implications for ecosystem service estimates. Ecology and Society 27(2):11.ABSTRACT
Bats provide a number of ecosystem services in agricultural areas, including the predation of night-flying insects, for which they are estimated to save agricultural industries billions of dollars per year. Intensive agriculture has many negative effects on biodiversity, and it is important to understand how wildlife exploit available habitats to allow persistence in these human-modified landscapes. To better evaluate the effectiveness of bats’ pest-controlling services, and to increase understanding of bat foraging behavior in these historically open grassland landscapes, we estimated bat activity and insect abundance in and around crop fields in southeast Nebraska, USA. Specifically, we used a novel acoustic grid sampling approach to document and visualize spatiotemporal activity patterns by different bat species over agricultural fields and forested habitat along crop field edges. Bat activity was highest in areas with the most forested edge habitat, and sites with more trees and water typically had more species present. Bat species and activity was low in isolated forest fragments and sites with minimal habitat edges, but overall insect volume did not decline away from field edges, suggesting that ecosystem services provided by bats likely diminish not because of a decline in resource availability, but because of the lack of structure. Woodland interfaces are important habitats for bats, and the invasion of grasslands by woody species in the Great Plains has increased available bat habitat, and therefore services provided by bats, but with a cost to grasslands and the ecological services they provide. However, although bats are clearly important insect predators that benefit agricultural activities, our ability to quantify the ecosystem services they provide will be greatly improved with a more nuanced understanding of how their activity varies relative to habitat structure and scale within the landscapes where these services are required.INTRODUCTION
Intensive agriculture is one of the primary drivers of global change, with rapid global expansion resulting in the destruction of natural ecosystems, as well as increased fertilizer and pesticide use (Matson et al. 1997, Tilman et al. 2001, Meehan et al. 2011). In many regions, including the midwestern United States, crops are planted in large landscape-scale monocultures to maximize production and profit, causing ecosystem simplification, species decline, and the loss of ecosystem services provided by wildlife (Daily et al. 2001, Tscharntke et al. 2012, De Beenhouwer et al. 2013).
As the primary predator of night flying insects, bats are thought to play a major role in controlling insect pest populations (Cleveland et al. 2006, Boyles et al. 2011, Maine and Boyles 2015) and serve as bio-indicators of ecosystem health (Park 2015). In rural landscapes, areas of forest cover are important habitat for tree-roosting bats, as well as both narrow and edge-space foraging species (Estrada and Coates-Estrada 2001, Henderson and Broders 2008, Wolcot and Volunec 2012). Isolated trees, riparian zones, and water bodies have also been indicated as important habitat for these flying mammals (Lumsden and Bennett 2005, Fuentes-Montemayor et al. 2013, Kalda et al. 2014), even in grassland habitats (Holloway and Barclay 2000). Bats will also navigate using vegetation structures as location references and guides while flying (Jensen 2005). Hedgerows, windbreaks, and other linear features are common in many agricultural landscapes and are utilized by many foraging bats (Frey-Ehrenbold et al. 2013, Kelm et al. 2014). Forests and streams can additionally serve as travel corridors.
In agricultural areas, not all bat species respond similarly to landscape modification. Some species forage more intensively over agricultural fields than in forest or other available non-crop habitat (Williams-Guillén and Perfecto 2011, Noer et al. 2012, Heim et al. 2016). Fast flying bats have been documented foraging in a variety of habitats (Estrada and Coates-Estrada 2002), whereas those with more specialized diets and differing wing morphologies seem less common in fragmented, intensively farmed areas, suggesting an unwillingness or inability to move through open habitats (Walsh and Harris 1996, Harvey and González Villalobos 2007, Freudmann et al. 2015). Agricultural intensification has also been shown to have minimal influence on species richness in some regions (Williams-Guillén and Perfecto 2011, Treitler et al. 2016, Mtsetfwa et al. 2018), in contrast to other findings documenting negative effects on forest specialists (Henderson and Broders 2008, Cleary et al. 2016, Heim et al. 2016).
In agricultural landscapes, bats likely play important roles in insect pest control and suppression, and attempts have been made to quantify these services (Kunz et al. 2011, Maine and Boyles 2015). Cleveland et al. (2006) investigated Brazilian free-tailed bat (Tadarida brasiliensis) and cotton bollworm (Helicoverpa armigera) behavioral interactions across cotton plantations in eight southwest Texas counties, estimating the bats pest control services at roughly $183/hectare. Boyles et al. (2011) applied these ecosystem service estimates to the cropland area of the continental United States, arriving at an approximate value range of $3.7 to $53 billion/year. Maine and Boyles (2015) also extrapolated their findings from a few corn fields in Illinois to global corn savings of approximately U.S. $1 billion/year. However, simply scaling up results from one or a few species in a handful of study sites fails to account for habitat requirements, geographic ranges, and foraging behaviors of the different bat species that actually occupy the areas extrapolated to. This strategy also neglects the crucial concept of the scale with which species interact with their environment (Peterson et al. 1998, Hein et al. 2006, Treitler et al. 2016).
The purpose of this study was to examine and visualize the spatial activity patterns of bat species in intensively managed agricultural fields of corn and soybean in the midwestern United States using a novel acoustic sampling method. Determining how bat species utilize the current agricultural matrix is critical in quantifying their roles in pest suppression, identifying habitat needs for conservation, and gaining a better understanding of bat foraging behavior in these landscapes. Our objectives were the following: (1) determine if bat species forage homogenously over crop fields or are limited to certain habitat features, and (2) determine if insect biomass is higher along field edge habitats than in crop fields. We hypothesized that bat activity would be different for each species and not homogenous, but rather concentrated by areas with woody vegetation and water, with little activity in open crop fields, and that insect biomass would decline away from edges. The spatial extents of bat foraging distribution over time through this unique sampling method presents a different way to view bat movements, and offers a deeper insight into bat ecology, with considerable implications for the economic value in ecosystem services they provide.
METHODS
Study area
We conducted this study on privately managed crop fields in Gage and Lancaster Counties, located in southeast Nebraska. Part of the North American Great Plains region, Nebraska was historically a prairie ecosystem dominated landscape prior to European settlement. The area was heavily managed for the production of corn and soybean during our study, and was characterized by large tracts of open farmland on flat upland plains interspersed with riparian buffers, windbreaks, and patches of mature lowland forest (Fig. 1).
Site selection and detector setup
We selected 11 study sites using ArcMap (10.7.1), each encompassing an area of 400 m by 600 m of corn and/or soybean, as well as one restored native tallgrass prairie site (Fig. 1). Sites were on average 5.36 km apart (minimum 0.83 km, maximum 39.01 km) and were all at least partially bordered by some form of tree cover or water source, except two sites with no immediate habitat edge. Agricultural sites were privately owned and managed by different landowners, whereas the restored prairie was maintained by the National Park Service. We used AnaBat Express passive zero cross acoustic detectors (Titley Scientific, Brendale, Australia; https://www.titley-scientific.com/us/) to record bat activity. Detectors were set to a 100 percent recording rate and triggered by echolocation calls above 8 kHz, which were recorded in zero-cross format. Because each Express unit has a maximum optimal range of 50 m, we used ArcGIS to create a layer of points each 100 m apart to form a transect grid that bordered the habitat feature and extended into the open crop field (Fig. 1), spanning a detection area 400 m by 600 m that employed 24 detectors (Mtsetfwa et al. 2018). We entered the coordinates for these locations into hand-held Garmin GPSMAP 64 units for actual detector placement in the field.
We mounted each detector on a modified painter’s pole extended 4 m above the ground so all units cleared any crop cover for increased recording quality (Mtsetfwa et al. 2018). We positioned detectors such that every detector’s omnidirectional microphone faced into the open and away from tree clutter. We left detectors at each deployment site for four consecutive nights, with each detector set to begin recording from at least 30 minutes before sunset until 30 minutes after sunrise. If there was sustained heavy rain, low temperatures, or winds greater than Beaufort Number 5 during a deployment session, we left detectors out for additional nights as needed, until four sampling nights were obtained. Issues with battery life occurred on two sites, so we immediately redeployed the few problem detectors and used a combination of different nights in both cases, using as many identical nights as possible across detectors. Sampling took place during summer, when Nebraska bats are most active, from June through August in 2019. We sampled sites in succession, although some survey dates overlapped.
Acoustic analysis
We downloaded and ran raw acoustic detector files through AnalookW software (Titley Scientific; Mancina et al. 2012, Fuentes-Montemayor et al. 2013) to convert files to individual bat calls, i.e., a sequence of at least two echolocation pulses, for further analysis. We analyzed all recordings using Kaleidoscope v5.1.9 with the Bats of North America 5.1.0 classifier set to “+1 More Accurate” to minimize the number of misidentifications (Wildlife Acoustics, Concord, Massachusetts, USA; https://www.wildlifeacoustics.com/). We used Kaleidoscope’s recommended default signal parameters as follows: 8–120 kHz frequency range, 2–500 ms length of detected pulses, 500 ms inter-syllable gap, 2 ms minimum number of pulses, and advanced signal processing feature activated, which enhances signals for cleaner output. Under the Bats of North America 5.1.0 classifier, the selected species included in the identification process were: big brown bat (Eptesicus fuscus), eastern red bat (Lasiurus borealis), hoary bat (Lasiurus cinereus), silver-haired bat (Lasionycteris noctivagans), little brown myotis (Myotis lucifugus), northern myotis (Myotis septentrionalis), evening bat (Nycticeius humeralis), and tricolored bat (Perimyotis subflavus; Czaplewski et al. 1979). Kaleidoscope Pro looks for unique characteristics in the acoustic calls that makes the identification process of calls to species more feasible, and also outputs the presence probability of a particular species (Willcox et al. 2017). By summing up the numbers of identified bat call recordings, we obtained an index of bat species activity by location and time.
Interpolation maps
We analyzed all identification output results from Kaleidoscope Pro in R (3.6.2; R Core Team 2019). Additionally, we created spatial interpolation maps for each site using the ArcMap (10.7.1) kriging tool method, which relies on spatial autocorrelation to assist in estimating the degree of bat activity in areas between detectors. We matched species recordings to each detector point making up the grid shapefile, and used these to create bins and an interpolated surface map layer. Because bat data were not normally distributed, we used simple kriging with normal score transformation, which transforms the data to standard normal distribution. We fit the stable semivariogram model provided by ArcMap to the data, and also performed a sensitivity test between several transformation methods, including log, box-cox, none, and normal score, and found no significant differences between predicted and expected point values among the models in the mapping results. For kriging results, we classified contours using equal intervals, with the number of classes relative to the number of calls for each species. Bins for highly active bats were in roughly 20 call increments, with higher bins averaging an additional five call recordings per night to denote a “significant” activity level increase. Less active species, however, especially those with little spatial autocorrelation, we grouped in just enough classes to show differences in the interpolated surface. We did not create maps for species at sites with low numbers of recordings (less than 10 calls over the four-night period) or little spatial autocorrelation (Moran’s Index ≤ 0.01), which kriging could not support. For active species we also created interpolation maps using the same method but in half-hour increments to visualize species spatiotemporal movements within sampling areas.
Insect analysis
To gain insight into insect prey distribution, we collected insects at increasing distances from tree cover at six of the 11 plots, on account of time and personnel constraints. We collected insects with 22 W fluorescent black light traps (Black Light Trap 2851M, BioQuip Products, Rancho Dominguez, California, USA) using three traps for each sampling session, placed at 20 m, 220 m, and 420 m from the field edge. We conducted insect sampling on subsequent nights within 30 days after the acoustic component had taken place, during similar weather conditions as bat surveys, and after all agricultural sites had been treated with pesticides. We suspended each light trap just above crop canopy by running a rope cord through the trap top handle and tied to extendable painter’s poles on either side of the trap, which could be adjusted as needed for the trap light to clear the crop canopy. Insects then drawn to the light fell through a funnel into a bucket containing a dichlorvos fumigant strip. We turned on traps within 45 minutes of sunset and collected them after 5 hours of sampling. We emptied light trap contents into Ziploc bags for freezer storage and upon analysis allowed approximately 24 hours to thaw. To estimate volume, we transferred insects from each trap separately to 250 mL graduated cylinders and settled by tapping prior to the measurement of volume (Allen et al. 2001). This provides an index of insect biomass at different distances from field edges to determine if insect biomass decays with distance from tree cover.
Statistical analysis
We conducted all analyses in R (3.6.2, R Core Team 2019) and pooled data from four nights at each site, given that bat activity has been found to vary even on a nightly basis relative to variables including temperature, prey, weather, and moon phase (Kunz 1973, Hayes 1997, Ciechanowski et al. 2008, Barros et al. 2014, Heim et al. 2017). We constructed a generalized linear mixed model to determine the influence of distance from tree cover on levels of bat activity, with distance and site as fixed and random factors, respectively. We used total bat recordings by each detector as the response variable, and fitted the model using a negative binomial distribution (log-likelihood: -5475; AIC: 10958) over Poisson (log-likelihood: -41234; AIC: 82491). We also created boxplots of species activity from raw data, grouping calls by detector locations within 25, 50, 75, 100, 150, 200, 300, 400, 500, and over 600 m from tree cover to record the effect of woody habitats on bat activity over crops. For three sites where detector grids were not bordered directly by any significant landscape feature, distances were measured to the nearest source of tree cover in ArcMap (10.7.1), which in most cases were small shelterbelts and isolated trees, all within 832 m.
RESULTS
Bat activity
Total bat activity was higher at the six sites with the most tree cover and water availability (median = 153 recordings, Q1 = 0, Q2 = 18.75, Q3 = 978.25, Q4 = 4987), than at the five sites with little to no edge habitat (median = 40 recordings, Q1 = 0, Q2 = 3, Q3 = 93.75, Q4 = 841). Hoary bats (11,842 recordings) and big brown bats (10,888 recordings) were the most frequently detected, whereas evening (7259 recordings), eastern red (6105 recordings), and silver-haired bats (1466 recordings) were also recorded at all sites. Additionally, northern long-eared bats (159 recordings) and little brown bats (153 recordings) were active on a few sites where larger forest fragments and tree cover were present (Fig. 2a). Tricolored bats (131 recordings) were similar to both Myotis species with activity on forested sites, but in one instance were also detected over a corn field with virtually no tree cover (Fig. 2b). Big brown bats were most active at sites with water present, as were eastern red, hoary, silver-haired, and evening bats (Fig. 2a). However, big brown, eastern red, hoary, and evening bats were also regularly present in areas of little tree cover as well (Fig. 2b).
Interpolation maps
Spatial maps for most sites revealed overall bat activity to be highly concentrated along woody edges and over small water bodies, and rapidly decline into open crop fields (Figs. 3–9). Sites with minimal landscape features surrounding the sample fields tended to have little to no spatial autocorrelation across samples and species (Moran’s Index ≤ 0.3). In these instances, the semivariogram kriging model consisted of a very shallow or flat line, and interpolation maps were not applicable. Thus, we did not map bat activity for four sites. Interpolated maps over time revealed species across sites seemed to expand their flight activities near sunset and sunrise, and largely confined movements to wooded areas or water bodies throughout the remainder of each night (Figs. 10–21).
Our model explained some of the data variation (McFadden pseudo R² = 0.05) and indicated that bat activity was influenced by the proximity of trees and woodland areas (estimate = -0.0022, SE = 0.0004, z-value = -5.935, p-value = 0.0001). When constructing boxplots (Fig. 2), we plotted along a log-scale y axis to include the furthest outlier at treed sites (Fig. 2a) within 25 m for each species (recordings: big brown 6116, hoary 3477, evening 1753, eastern red 1620, silver-haired 531, northern long-eared 122, little brown 83, tricolored 27). All bat species were most active within 200 m of edge habitats.
Insect sampling
Of the five agricultural sites sampled with insect light traps, insect volume overall did not change substantially with increasing distance from wooded areas (Fig. 22). Two soybean fields that bordered single windbreaks both had the highest insect volume totals of our sampled sites (726 mL, 646 mL), whereas the other three crop sites bordered creek (148 mL), woodland (605 mL), and buffer strip (542 mL), respectively. The restored tallgrass prairie site had the second lowest total insect volume of our sites (232 mL) and bordered a riparian woodland.
DISCUSSION
This is the first study to visualize the spatiotemporal habitat use by bats in an agricultural landscape utilizing acoustic grids. Results based on acoustic data indicated that forested areas, edge habitats, and patches of tree cover are important areas for bats in a typical Great Plains agricultural landscape. Bat activity over crop fields was lower than in forested edges and decreased with increasing distance from these edge types. Additionally, bat activity seemingly only occurred in airspace above crop fields for a small subset of the total night hours. The apparent infrequency of bat activity over open farmland, despite available insects, may have considerable implications for the ecosystem services bats may actually provide, as opposed to the economic values projected by prior small-scale extrapolative studies.
Although bat activity was highest along forested areas, a few bat species were still present at even the most isolated features sampled, including tree patches, single trees, windbreaks, and open field detectors 800 m away from woody vegetation. This behavior demonstrates that even remote and small-scale resources are important refuges for wildlife, even in large-scale agriculturally converted landscapes. Other studies have similarly found bats foraging by scattered and isolated trees (Lumsden and Bennett 2005, Le Roux et al. 2018). In this study, the most common species detected at such open and remote locations were often hoary, big brown, evening, and eastern red bats, which suggests that these species venture out into open habitats. Hoary and eastern red bats are regarded as fast flying aerial hawkers, often found in open spaces by vegetation and sometimes at high altitude (Norberg and Rayner 1987). Big brown, silver-haired, and evening bats will also hawk prey around cluttered and open areas (Norberg and Rayner 1987, Jones et al. 2016). Although in this study, overall activity at these remote locations was very low, such features could serve as stepping stones and foraging opportunities as bats move through the landscape, and might also serve to draw bats further from habitat edges into open spaces (Estrada and Coates-Estrada 2001, Heim et al. 2018). In this study, bat use on the prairie site did not differ much from distributions on crop plots, suggesting that prior to European settlement, bat prairie use was likely restricted mostly to riparian areas and isolated trees (Holloway and Barclay 2000, Heim et al. 2015, Treitler et al. 2016), and by fewer species (Benedict 2004, Anderson et al. 2017). Because some bats are capable of flying over long distances through open landscapes (Estrada and Coates-Estrada 2002), the low number of echolocation recordings at exposed or isolated tree cover sites could indicate bats are simply passing through and not necessarily foraging, or are flying at higher altitudes (Griffin and Thompson 1982, Fenton and Griffin 1997, Frick et al. 2012) in these areas. Some bats can change foraging behavior to take advantage of sudden insect abundances (Charbonnier et al. 2014, Maine and Boyles 2015), even in open habitat (Müller et al. 2012), and can track migrating insect populations (McCracken et al. 2012).
We obtained limited data on little brown, northern long-eared, and tricolored bats. We detected these three species almost exclusively at sites containing large amounts of tree cover, and they were most active in forest interior and edge habitats. These species are regarded as clutter-adapted species, capable of hawking, hover-gleaning, and trawling insects from vegetation, ground, and water surfaces (Norberg and Rayner 1987, Ratcliffe and Dawson 2003, Jones et al. 2016). In North America, they are among the most impacted by white-nose syndrome (Frick et al. 2015). Despite this, other studies have found that even these clutter-adapted species will forage in open areas over forested ones, likely in instances where the overall fragmented landscape complements roosting and foraging sites (Ethier and Fahrig, 2011, Monck-Whipp et al. 2018).
Unlike bat activity, insect volume collected from light traps was not necessarily highest at the habitat edge (Fig. 22) and did not show a single, systematic pattern in relation to distance from forests or field edges. Thus, the spatial patterning of bat activities may not be driven by the location of insect prey. Flying insects in these agricultural environments tend to be affected by wind and daily weather conditions (Lewis 1969, Grüebler et al. 2008) and field distributions can vary by species (Dix et al. 1995). Similarly, inclement weather can also alter and reduce bat activity (Erickson 2002, Parsons 2003, Frick et al. 2012). Nevertheless, our results indicate that although overall insect volume remained largely unchanged over crop fields from tree cover, bat foraging activity primarily occurs along habitat edges with seemingly very little to no activity over crop fields. Even those bats that did venture away from wooded areas seemingly did so for only a mere fraction of their nightly movements (Fig. 10–21). Consequently, insects and crop pests that remain in open tracts of agricultural crops or along isolated tree patches are far less likely to be consumed by bat species in the area, and can continue to inflict agricultural damages. Although insects were not identified in this study, insect abundance has been found to increase with higher land-use intensity, although accompanied by a decrease in insect size and diversity (Treitler et al. 2016) and a likely reduction in preferred prey species by different bats. It is also possible that when we sampled some fields for insects almost a month after bat sampling, insect abundance or composition may have changed significantly and would not accurately reflect prior bat behavior at those individual sites.
Bats under laboratory conditions have demonstrated an ability to consume hundreds of insects at an hourly rate (Griffin et al. 1960, Coutts et al. 1973). Although this rate of consumption is likely not realistic in the wild, some species are capable of eating close to their own body weight each night (Charles-Dominique 1991, Kunz et al. 1995), and bats are thought to play a major role in controlling insect pest populations. Studies suggest bats can save farmers billions of dollars per year in crop damage reductions (Cleveland et al. 2006, Boyles et al. 2011, Maine and Boyles 2015). However, these are national or even global extrapolations based on a few bat species in small-scale studies on a single crop, or in the case of Boyles et al. (2011), the Brazilian free-tailed bat, an abundant generalist species that locally forms the largest concentrations of mammals on earth (McCracken 2003) but does not inhabit most of North America. Such extrapolations assume all bats use all crop areas equally, however many species are habitat specialized and appear to primarily capitalize on areas with tree cover. Consequently, these bats are not likely to spend considerable time consuming insects in expansive tracts of open crop monocultures. This illustrates the importance of scale, landscape connectivity, and how different species interact with their surroundings (Tscharntke et al. 2005, Ethier and Fahrig 2011, Treitler et al. 2016). Therefore, test results obtained from closed environments that exclude natural processes cannot always be applied to predict outcomes and behaviors in actual complex and dynamic ecosystems.
Our study may have considerable implications for the large-scale economic benefits that bats actually provide, relative to what has been suggested previously (Boyles et al. 2011). Regardless, there is little question that bats consume large numbers of insects and crop pests on a nightly basis, which undoubtedly mitigates potential insect damage to some degree. In intensively managed large-scale agricultural landscapes such as the midwestern United States, smaller crop field size, additional tree cover in the forms of windbreaks and shelterbelts, as well as ponds and riparian waterways would all serve to increase bat activity and the economic impact they could provide through insect consumption. Although advantageous for bats, however, tree species, if neglected or incorrectly managed, can also invade sensitive ecosystems like grasslands and prairies, which dominated states like Nebraska prior to European settlement and subsequent agricultural conversion (Leis et al. 2017). Consequently, it is important to focus on species-level habitat differences and the scale of those habitat associations in order to formulate systems-level resource management strategies.
CONCLUSION
Through a novel acoustic grid sampling approach we were able to visualize bat spatiotemporal activity patterns over agricultural fields. We found that the amount of bat activity, number of species, and spatial use in a typical Great Plains agricultural landscape is profoundly influenced by the presence of wooded areas and accessible water. Overall bat activity was concentrated over and alongside these areas, despite the availability of insects over crop fields. Additionally, although areas of tree cover are advantageous for bats, woody encroachment is also detrimental to grassland ecosystems (Donovan et al. 2018) and can even result in a loss of ecosystem services. As the first study to visualize bat spatiotemporal movements, these findings have considerable implications for the estimated economic value of the ecosystem services that bats actually provide, given that estimations by prior studies disregard the scale of individual species habitat associations.
RESPONSES TO THIS ARTICLE
Responses to this article are invited. If accepted for publication, your response will be hyperlinked to the article. To submit a response, follow this link. To read responses already accepted, follow this link.ACKNOWLEDGMENTS
This study is based upon work supported by the National Science Foundation under Grant No. DGE-1735362. I thank D. Uden for additional guidance in spatial analyses methods, Homestead National Monument for their support of the project, and A. Oetting for assisting in data collection throughout the entire project.
DATA AVAILABILITY
The data and code that support the findings of this study are available on request from the corresponding author, CF.
LITERATURE CITED
Allen, C. R., R. S. Lutz, T. Lockley, S. A. Phillips Jr., and S. Demarais. 2001. The non-indigenous ant, Solenopsis invicta, reduces loggerhead shrike and native insect abundance. Journal of Agricultural and Urban Entomology 18(4):249-259. https://www.researchgate.net/publication/286614167
Anderson, B. R., K. Geluso, H. W. Otto, and L. Bishop-Boros. 2017. Westward expansion of the evening bat (Nycticeius humeralis) in the United States, with notes on the first record from New Mexico. Western North American Naturalist 77(2):223-229. https://doi.org/10.3398/064.077.0210
Barros, M. A. S., D. M. A. Pessoa, and A. M. Rui. 2014. Habitat use and seasonal activity of insectivorous bats (Mammalia: Chiroptera) in the grasslands of southern Brazil. Zoologia 31(2):153-161. https://doi.org/10.1590/S1984-46702014000200006
Benedict, R. A. 2004. Reproductive activity and distribution of bats in Nebraska. Western North American Naturalist 64(2):231-248. https://scholarsarchive.byu.edu/wnan/vol64/iss2/10
Boyles, J. G., P. M. Cryan, G. F. Mccracken, and T. H. Kunz. 2011. Economic importance of bats in agriculture. Science 332(6025):41-42. https://doi.org/10.1126/science.1201366
Charbonnier, Y., L. Barbaro, A. Theillout, and H. Jactel. 2014. Numerical and functional responses of forest bats to a major insect pest in pine plantations. PLoS ONE 9(10):e109488. https://doi.org/10.1371/journal.pone.0109488
Charles-Dominique, P. 1991. Feeding strategy and activity budget of the frugivorous bat Carollia perspicillata (Chiroptera: Phyllostomidae) in French Guiana. Journal of Tropical Ecology 7(2):243-256. https://doi.org/10.1017/S026646740000540X
Ciechanowski, M., T. Zając, A. Biłas, and R. Dunajski. 2008. Spatiotemporal variation in activity of bat species differing in hunting tactics: effects of weather, moonlight, food abundance, and structural clutter. Canadian Journal of Zoology 85:1249-1263. https://doi.org/10.1139/Z07-090
Cleary, K. A., L. P. Waits, and B. Finegan. 2016. Agricultural intensification alters bat assemblage composition and abundance in a dynamic Neotropical landscape. Biotropica 48(5):667-676. https://doi.org/10.1111/btp.12327
Cleveland, C. J., M. Betk, P. Federico, J. D. Frank, T. G. Hallam, J. Horn, J. D. López Jr., G. McCracken, R. Medellin, A. Moreno-Valdez, C. Sansone, J. Westbrook, and T. Kunz. 2006. Estimation of the economic value of the pest control service provided by the brazilian free-tailed bat in the winter garden region of south-central Texas. Frontiers in Ecology and the Environment 4(5):238-243. https://doi.org/10.1890/1540-9295(2006)004[0238:EVOTPC]2.0.CO;2
Coutts, R., M. Fenton, and E. Glen. 1973. Food intake by captive Myotis lucifugus and Eptesicus fuscus (Chiroptera: Vespertilionidae). Journal of Mammalogy 54(4):985-990. https://doi.org/10.2307/1379098
Czaplewski, N. J., J. P. Farney, J. K. Jones Jr., and J. D. Druecker. Synopsis of bats of Nebraska. Occasional Papers, The Museum, Texas Tech University 61:1-24.
Daily, G. C., P. R. Ehrlich, and G. Arturo Sanchez-Azofeifa. 2001. Countryside biogeography: use of human-dominated habitats by the avifauna of southern Costa Rica. Ecological Applications 11(1):1-13. https://doi.org/10.1890/1051-0761(2001)011[0001:CBUOHD]2.0.CO;2
Davidai, N., J. K. Westbrook, J. P. Lessard, T. G. Hallam, and G. F. McCracken. 2015. The importance of natural habitats to Brazilian free-tailed bats in intensive agricultural landscapes in the winter garden region of Texas, United States. Biological Conservation 190:107-114. https://doi.org/10.1016/j.biocon.2015.05.015
De Beenhouwer, M., R. Aerts, and O. Honnay. 2013. A global meta-analysis of the biodiversity and ecosystem service benefits of coffee and cacao agroforestry. Agriculture, Ecosystems & Environment 175:1-7. https://doi.org/10.1016/j.agee.2013.05.003
Dix, M. E., R. J. Johnson, M. O. Harrell, R. M. Case, R. J. Wright, L. Hodges, J. R. Brandle, M. M. Schoeneberger, N. J. Sunderman, R. L. Fitzmaurice, L. J. Young, and K. G. Hubbard. 1995. Influences of trees on abundance of natural enemies of insect pests: a review. Agroforestry Systems 29:303-311. https://doi.org/10.1007/BF00704876
Donovan, V. M., J. L. Burnett, C. H. Bielski, H. E. Birgé, R. Bevans, D. Twidwell, and C. R. Allen. 2018. Social-ecological landscape patterns predict woody encroachment from native tree plantings in a temperate grassland. Ecology and Evolution 8(19):9624-9632. https://doi.org/10.1002/ece3.4340
Erickson, J. L., and S. D. West. 2002. The influence of regional climate and nightly weather conditions on activity patterns of insectivorous bats. Acta Chiropterologica 4(1):17-24. https://doi.org/10.3161/001.004.0103
Estrada, A., and R. Coates-Estrada. 2001. Bat species richness in live fences and in corridors of residual rain forest vegetation at Los Tuxtlas, Mexico. Ecography 24(1):94-102. https://doi.org/10.1034/j.1600-0587.2001.240111.x
Estrada, A., and R. Coates-Estrada. 2002. Bats in continuous forest, forest fragments and in an agricultural mosaic habitat-island at Los Tuxtlas, Mexico. Biological Conservation 103(2):237-245. https://doi.org/10.1016/S0006-3207(01)00135-5
Ethier, K., and L. Fahrig. 2011. Positive effects of forst fragmentation, independent of forest amount, on bat abundance in eastern Ontario, Canada. Landscape Ecology 26:865-876. https://doi.org/10.1007/s10980-011-9614-2
Fenton, M. B., and D. R. Griffin. 1997. High-altitude pursuit of insects by echolocating bats. Journal of Mammalogy 78(1):247-250. https://doi.org/10.2307/1382658
Freudmann, A., P. Mollik, M. Tschapka, and C. H. Schulze. 2015. Impacts of oil palm agriculture on phyllostomid bat assemblages. Biodiversity and Conservation 24:3583-3599. https://doi.org/10.1007/s10531-015-1021-6
Frey-Ehrenbold, A., F. Bontadina, R. Arlettaz, and M. K. Obrist. 2013. Landscape connectivity, habitat structure and activity of bat guilds in farmland-dominated matrices. Journal of Applied Ecology 50(1):252-261. https://doi.org/10.1111/1365-2664.12034
Frick, W. F., S. J. Puechmaille, J. R. Hoyt, B. A. Nickel, K. E. Langwig, J. T. Foster, K. E. Barlow, T. Bartonička, D. Feller, A. J. Haarsma, C. Herzog, I. Horáček, J. van der Kooij, B. Mulkens, B. Petrov, R. Reynolds, L. Rodrigues, C. W. Stihler, G. G. Turner, and A. M. Kilpatrick. 2015. Disease alters macroecological patterns of North American bats. Global Ecology and Biogeography 24(7):741-749. https://doi.org/10.1111/geb.12290
Frick, W. F., P. M. Stepanian, J. F. Kelly, K. W. Howard, C. M. Kuster, T. H. Kunz, and P. B. Chilson. 2012. Climate and weather impact timing of emergence of bats. PLoS ONE 7(8):e42737. https://doi.org/10.1371/journal.pone.0042737
Fuentes-Montemayor, E., D. Goulson, L. Cavin, J. M. Wallace, and K. J. Park. 2013. Fragmented woodlands in agricultural landscapes: the influence of woodland character and landscape context on bats and their insect prey. Agriculture, Ecosystems & Environment 172:6-15. https://doi.org/10.1016/j.agee.2013.03.019
Griffin, D. R., and D. Thompson. 1982. High altitude echolocation of insects by bats. Behavioral Ecology and Sociobiology 10:303-306. https://doi.org/10.1007/BF00302821
Griffin, D. R., F. A. Webster, and C. R. Michael. 1960. The echolocation of flying insects by bats. Animal Behavior 8(3-4):141-154. https://doi.org/10.1016/0003-3472(60)90022-1
Grüebler, M. U., M. Morand, and B. Naef-Daenzer. 2008. A predictive model of the density of airborne insects in agricultural environments. Agriculture, Ecosystems & Environment 123(1-3):75-80.
Harvey, C. A., and J. A. González Villalobos. 2007. Agroforestry systems conserve species-rich but modified assemblages of tropical birds and bats. Biodiversity and Conservation 16:2257-2292. https://doi.org/10.1007/s10531-007-9194-2
Hayes, J. P. 1997. Temporal variation in activity of bats and the design of echolocation-monitoring studies. Journal of Mammalogy 78(2):514-524. https://doi.org/10.2307/1382902
Heim, O., J. Lenski, J. Schulze, K. Jung, S. Kramer-Schadt, J. A. Eccard, and C. C. Voigt. 2018. The relevance of vegetation structures and small water bodies for bats foraging above farmland. Basic and Applied Ecology 27:9-19. https://doi.org/10.1016/j.baae.2017.12.001
Heim, O., L. Lorenz, S. Kramer-Schadt, K. Jung, C. C. Voigt, and J. Eccard. 2017. Landscape and scale-dependent spatial niches of bats foraging above intensively used arable fields. Ecological Processes 6:24. https://doi.org/10.1186/s13717-017-0091-7
Heim, O., A. Schroder, J. Eccard, K. Jung, and C. C. Voigt. 2016. Seasonal activity patterns of European bats above intensively used farmland. Agriculture, Ecosystems & Environment 233:130-139 https://doi.org/10.1016/j.agee.2016.09.002
Heim, O., J. T. Treitler, M. Tschapka, M. Knornschild, and K. Jung. 2015. The importance of landscape elements for bat activity and species richness in agricultural areas. PLoS ONE 10(7):e0134443. https://doi.org/10.1371/journal.pone.0134443
Hein, L., K. van Koppen, R. S. de Groot, and E. C. van Ierland. 2006. Spatial scales, stakeholders and the valuation of ecosystem services. Ecological Economics 57(2):209-228. https://doi.org/10.1016/j.ecolecon.2005.04.005
Henderson, L. E., and H. G. Broders. 2008. Movements and resource selection of the northern long-eared myotis (Myotis septentrionalis) in a forest-agriculture landscape. Journal of Mammalogy 89(4):952-963. https://doi.org/10.1644/07-MAMM-A-214.1
Holloway, G. L., and R. M. R. Barclay. 2000. Importance of prairie riparian zones to bats in southeastern Alberta. Écoscience 7(2):115-122. https://doi.org/10.1080/11956860.2000.11682579
Jensen, M. E. 2005. Echolocating bats can use acoustic landmarks for spatial orientation. Journal of Experimental Biology 208(23):4399-4410. https://doi.org/10.1242/jeb.01901
Jones, P. L., R. A. Page, and J. M. Ratcliffe. 2016. To scream or to listen? Prey detection and discrimination in animal-eating bats. Pages 93-116 in M. Fenton, A. Grinnell, A. Popper, and R. Fay, editors. Bat bioacoustics. Springer Handbook of Auditory Research, vol 54. Springer, New York, New York, USA. https://doi.org/10.1007/978-1-4939-3527-7_4
Kalda, O., R. Kalda, and J. Liira. 2014. Multi-scale ecology of insectivorous bats in agricultural landscapes. Agriculture, Ecosystems & Environment 199:105-113. https://doi.org/10.1016/j.agee.2014.08.028
Kelm, D. H., J. Lenski, V. Kelm, U. Toelch, and F. Dziock. 2014. Seasonal bat activity in relation to distance to hedgerows in an agricultural landscape in central Europe and implications for wind energy development. Acta Chiropterologica 16(1):65-73. https://doi.org/10.3161/150811014X683273
Kunz, T. H. 1973. Resource utilization: temporal and spatial components of bat activity in central Iowa. Journal of Mammalogy 54:14-32. https://doi.org/10.2307/1378869
Kunz, T. H., E. Braun de Torrez, D. Bauer, T. Lobova, and T. H. Fleming. 2011. Ecosystem services provided by bats. Annals of the New York Academy of Sciences 1223(1):1-38. https://doi.org/10.1111/j.1749-6632.2011.06004.x
Kunz, T. H., J. O. Whitaker Jr., and M. D. Wadanoli. 1995. Dietary energetics of the insectivorous Mexican free-tailed bat (Tadarida brasiliensis) during pregnancy and lactation. Oecologia 101:407-415. https://doi.org/10.1007/BF00329419
Leis, S. A., C. E. Blocksome, D. Twidwell, S. D. Fuhlendorf, J. M. Briggs, and L. D. Sanders. 2017. Juniper invasions in grasslands: research needs and intervention strategies. Rangelands 39(2):64-72. https://doi.org/10.1016/j.rala.2017.03.002
Le Roux, D. S., K. Ikin, D. B. Lindenmayer, A. D. Manning, and P. Gibbons. 2018. The value of scattered trees for wildlife: contrasting effects of landscape context and tree size. Diversity and Distributions 24(1):69-81. https://doi.org/10.1111/ddi.12658
Lewis, T. 1969. The distribution of flying insects near a low hedgerow. Journal of Applied Ecology 6(3):443-452. https://doi.org/10.2307/2401510
Lumsden, L. F., and A. F. Bennett. 2005. Scattered trees in rural landscapes: foraging habitat for insectivorous bats in south-eastern Australia. Biological Conservation 122(2):205-222. https://doi.org/10.1016/j.biocon.2004.07.006
Maine, J. J., and J. G. Boyles. 2015. Bats initiate vital agroecological interactions in corn. PNAS 112(40):12438-12443. https://doi.org/10.1073/pnas.1505413112
Mancina, C. A., L. García-Rivera, and B. W. Miller. 2012. Wing morphology, echolocation, and resource partitioning in syntopic Cuban mormoopid bats. Journal of Mammalogy 93(5):1308-1317. https://doi.org/10.1644/11-MAMM-A-331.1
Matson, P. A., W. J. Parton, A. G. Power, and M. J. Swift. 1997. Agricultural intensification and ecosystem properties. Science 277(5325):504-509. https://doi.org/10.1126/science.277.5325.504
McCracken, G. F. 2003. Estimates of population sizes in summer colonies of Brazilian free-tailed bats (Tadarida brasiliensis). Pages 21-30 in T. J. O’Shea and M. A. Bogans, editors. Monitoring trends in bat populations of the U.S. and Territories: problems and prospects. United States Geological Survey, Biological Resources Discipline, Information and Technology Report, USGS/BRD/IRT-2003-003. U.S. Geological Survey, Reston, Virginia, USA. https://pubs.usgs.gov/itr/2003/0003/report.pdf
McCracken, G. F., J. K. Westbrook, V. A. Brown, M. Eldridge, P. Federico, and T. H. Kunz. 2012. Bats track and exploit changes in insect pest populations. PLoS ONE 7(8):e43839. https://doi.org/10.1371/journal.pone.0043839
Meehan, T. D., B. P. Werling, D. A. Landis, and C. Gratton. 2011. Agricultural landscape simplification and insecticide use in the Midwestern United States. PNAS 108(28):11500-11505. https://doi.org/10.1073/pnas.1100751108
Monck-Whipp, L., A. E. Martin, C. M. Francis, and L. Fahrig. 2018. Farmland heterogeneity benefits bats in agricultural landscapes. Agriculture, Ecosystems & Environment 253:131-139. https://doi.org/10.1016/j.agee.2017.11.001
Mtsetfwa, F., R. A. McCleery, and A. Monadjem. 2018. Changes in bat community composition and activity patterns across a conservation-agriculture boundary. African Zoology 53(3):99-106. https://doi.org/10.1080/15627020.2018.1531726
Müller, J., M. Mehr, C. Bässler, M. B. Fenton, T. Hothorn, H. Pretzsch, H. J. Klemmt, and R. Brandl. 2012. Aggregative response in bats: prey abundance versus habitat. Oecologia 169:673-684. https://doi.org/10.1007/s00442-011-2247-y
Noer, C. L., T. Dabelsteen, K. Bohmann, and A. Monadjem. 2012. Molossid bats in an African agro-ecosystem select sugarcane fields as foraging habitat. African Zoology 47(1):1-11. https://doi.org/10.3377/004.047.0120
Norberg, U. M., and J. M. V. Rayner. 1987. Ecological morphology and flight in bats (Mammalia; Chiroptera): wing adaptations, flight performance, foraging strategy and echolocation. Philosophical Transactions of the Royal Society B Biological Sciences 316(1179):335-427. https://doi.org/10.1098/rstb.1987.0030
Park, K. J. 2015. Mitigating the impacts of agriculture on biodiversity: bats and their potential role as bioindicators. Mammalian Biology 80(3):191-204. https://doi.org/10.1016/j.mambio.2014.10.004
Parsons, K. N., G. Jones, and F. Greenaway. 2003. Swarming activity of temerate zone microchiropteran bats: effects of season, time of night and weather conditions. Journal of Zoology 261(3):257-267. https://doi.org/10.1017/S0952836903004199
Peterson, G., C. R. Allen, and C. S. Holling. 1998. Ecological resilience, biodiversity, and scale. Ecosystems 1:6-18. https://doi.org/10.1007/s100219900002
Ratcliffe, J. M., and J. W. Dawson. 2003. Behavioural flexibility: the little brown bat, Myotis lucifugus, and the northern long-eared bat, M. septentrionalis, both glean and hawk prey. Animal Behaviour 66(5):847-856. https://doi.org/10.1006/anbe.2003.2297
R Core Team. 2019. R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. https://www.R-project.org/
Tilman, D., J. Fargione, B. Wolff, C. D’Antonio, A. Dobson, R. Howarth, D. Schindler, W. H. Schlesinger, D. Simberloff, and D. Swackhamer. 2001. Forecasting agriculturally driven global environmental change. Science 292(5515):281-285. https://doi.org/10.1126/science.1057544
Treitler, J. T., O. Heim, M. Tschapka, and K. Jung. 2016. The effect of local land use and loss of forests on bats and nocturnal insects. Ecology and Evolution 6(13):4289-4297. https://doi.org/10.1002/ece3.2160
Tscharntke, T., Y. Clough, T. C. Wanger, L. Jackson, I. Motzke, I. Perfecto, J. Vandermeer, and A. Whitbread. 2012. Global food security, biodiversity conservation and the future of agricultural intensification. Biological Conservation 151(1):53-59. https://doi.org/10.1016/j.biocon.2012.01.068
Tscharntke, T., A. M. Klein, I. Steffan-Dewenter, and C. Thies. 2005. Landscape perspectives on agricultural intensification and biodiversity - ecosystem service management. Ecology Letters 8:857-874. https://doi.org/10.1111/j.1461-0248.2005.00782.x
Walsh, A. L., and S. Harris. 1996. Foraging habitat preferences of vespertilionid bats in Britain. Journal of Applied Ecology 33(3):508-518. https://doi.org/10.2307/2404980
Willcox, E. V., W. M. Giuliano, L. N. Watine, D. J. Mills, and M. G. Andreu. 2017. Forest structure and composition affect bats in a tropical evergreen broadleaf forest. Forests 8(9):317. https://doi.org/10.3390/f8090317
Williams-Guillén, K., and I. Perfecto. 2011. Ensemble composition and activity levels of insectivorous bats in response to management intensification in coffee agroforestry systems. PLoS ONE 6(1):e16502. https://journals.plos.org/plosone/article?id=10.1371/journal.pone.0016502
Wolcot, K. A., and K. Volunec. 2012. Bat activity at woodland/farmland interfaces in central Delaware. Northeastern Naturalist 19(1):87-98. https://doi.org/10.1656/045.019.0107
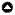
Fig. 1

Fig. 1. (A) Study area, with (B) sampling site locations and land cover types in Gage and Lancaster counties, Nebraska. All sites were corn or soybean fields, except one restored tallgrass prairie site. Each site was sampled using a grid of 24 acoustic detectors (C) with each detector placed 100 meters apart.
Fig. 2

Fig. 2. Boxplots of species activity levels with respect to tree cover proximity at sites with trees (A) and sites without trees (B). We plotted (A) on a log-scale y axis to include the highest outliers for each species (recordings: big brown 6116, hoary 3477, evening 1753, eastern red 1620, silver-haired 531, northern long-eared 122, little brown 83, tricolor 27), all of which occurred within 25 m of tree cover. We grouped recordings by detectors within 25 m, 50 m, 75 m, 100 m, 150 m, 200 m, 300 m, 400 m, 500 m, and over 600 m from tree cover.
Fig. 3

Fig. 3. Spatial interpolation results for bat species along a corn field in southeast Nebraska. Species detected with a Moran’s Index ≤ 0 include the eastern red (Lasiurus borealis; 2282 recordings), little brown (Myotis lucifugus; 22 recordings), and tricolored bat (Perimyotis subflavus; four recordings). Interpolation maps were made by kriging call results across four nights with a normal score transformation. Bins contain the bat call groupings. Site imagery taken 9 April 2020.
Fig. 4

Fig. 4. Spatial interpolation results for bat species along a corn field and pond in southeast Nebraska. Species detected with a Moran’s Index ≤ 0 include the silver-haired (Lasionycteris noctivagans; 12 recordings) and northern long-eared bats (Myotis septentrionalis; two recordings). Interpolation maps were made by kriging with a normal score transformation. Bins contain the bat call groupings. Site imagery taken 9 April 2020.
Fig. 5

Fig. 5. Spatial interpolation results for bat species along a soybean field in southeast Nebraska. Species detected with a Moran’s Index ≤ 0.12 include the hoary (Lasiurus cinereus; 77 recordings) and silver-haired bats (Lasionycteris noctivagans; seven recordings). Interpolation maps were made by kriging call results across four nights with a normal score transformation. Bins contain the bat call groupings. Site imagery taken 9 April 2020.
Fig. 6

Fig. 6. Spatial interpolation results for bat species along a corn and soybean field in southeast Nebraska. Species detected with a Moran’s Index ≤ 0 include the little brown (Myotis lucifugus; two recordings) and tricolored bat (Perimyotis subflavus; one recording). Interpolation maps were made by kriging call results across four nights with a normal score transformation. Bins contain the bat call groupings. Site imagery taken 9 April 2020.
Fig. 7

Fig. 7. Spatial interpolation results for bat species along a soybean and corn field in southeast Nebraska. Species detected with a weak spatial autocorrelation (Moran’s Index < 0.4) include the little brown (Myotis lucifugus; 14 recordings), northern long-eared (Myotis septentrionalis), and tricolored (Perimyotis subflavus) bats. Interpolation maps were made by kriging with a normal score transformation. Bins contain the bat call groupings. Site imagery taken 9 April 2020.
Fig. 8

Fig. 8. Spatial interpolation results for bat species along a corn field in southeast Nebraska. Bats detected with a Moran’s Index ≤ 0.12 include the northern long-eared (Myotis septentrionalis), hoary (Lasiurus cinereus), silver-haired (Lasionycteris noctivagans), little brown (Myotis lucifugus), and tricolored (Perimyotis subflavus) bats. Interpolation maps were made by kriging with a normal score transformation. Bins contain the bat call groupings. Site imagery taken 9 April 2020.
Fig. 9

Fig. 9. Spatial interpolation results for bat species along a tallgrass prairie in southeast Nebraska. Bats detected with weak autocorrelation (Moran’s Index ≤ 0.3) include eastern red (Lasiurus borealis), silver-haired (Lasionycteris noctivagans), little brown (Myotis lucifugus), northern long-eared (Myotis septentrionalis), and tricolored (Perimyotis subflavus) bats. Interpolation maps were made by kriging with a normal score transformation. Bins contain the bat call groupings. Site imagery taken 9 April 2020.
Fig. 10

Fig. 10. Spatiotemporal interpolation results for big brown bats (Eptesicus fuscus) along a wooded creek in southeast Nebraska. Maps were made by kriging with a normal score transformation with bins containing the call count groupings in half-hour increments spanning a period of four nights. Refer to Fig. 3 for still version.
Fig. 11

Fig. 11. Spatiotemporal interpolation results for hoary bats (Lasiurus cinereus) along a wooded creek in southeast Nebraska. Maps were made by kriging with a normal score transformation with bins containing the call count groupings in half-hour increments spanning a period of four nights. Refer to Fig. 3 for still version.
Fig. 12

Fig. 12. Spatiotemporal interpolation results for eastern red bats (Lasiurus borealis) along a corn field in southeast Nebraska. Maps were made by kriging with a normal score transformation with bins containing the call count groupings in half-hour increments spanning a period of four nights. Refer to Fig. 4 for still version.
Fig. 13

Fig. 13. Spatiotemporal interpolation results for big brown bats (Eptesicus fuscus) along a creek in southeast Nebraska. Maps were made by kriging with a normal score transformation with bins containing the call count groupings in half-hour increments spanning a period of four nights. Refer to Fig. 6 for still version.
Fig. 14

Fig. 14. Spatiotemporal interpolation results for eastern red bats (Lasiurus borealis) along a creek in southeast Nebraska. Maps were made by kriging with a normal score transformation with bins containing the call count groupings in half-hour increments spanning a period of four nights. Refer to Fig. 6 for still version.
Fig. 15

Fig. 15. Spatiotemporal interpolation results for hoary bats (Lasiurus cinereus) along a creek in southeast Nebraska. Maps were made by kriging with a normal score transformation with bins containing the call count groupings in half-hour increments spanning a period of four nights. Refer to Fig. 6 for still version.
Fig. 16

Fig. 16. Spatiotemporal interpolation results for big brown bats (Eptesicus fuscus) along a forest strip in southeast Nebraska. Maps were made by kriging with a normal score transformation with bins containing the call count groupings in half-hour increments spanning a period of four nights. Refer to Fig. 7 for still version.
Fig. 17

Fig. 17. Spatiotemporal interpolation results for eastern red bats (Lasiurus borealis) along a forest strip in southeast Nebraska. Maps were made by kriging with a normal score transformation with bins containing the call count groupings in half-hour increments spanning a period of four nights. Refer to Fig. 7 for still version.
Fig. 18

Fig. 18. Spatiotemporal interpolation results for hoary bats (Lasiurus cinereus) along a forest strip in southeast Nebraska. Maps were made by kriging with a normal score transformation with bins containing the call count groupings in half-hour increments spanning a period of four nights. Refer to Fig. 7 for still version.
Fig. 19

Fig. 19. Spatiotemporal interpolation results for evening bats (Nycticeius humeralis) along a forest strip in southeast Nebraska. Maps were made by kriging with a normal score transformation with bins containing the call count groupings in half-hour increments spanning a period of four nights. Refer to Fig. 7 for still version.
Fig. 20

Fig. 20. Spatiotemporal interpolation results for big brown bats (Eptesicus fuscus) along a forest fragment in southeast Nebraska. Maps were made by kriging with a normal score transformation with bins containing the call count groupings in half-hour increments spanning a period of four nights. Refer to Fig. 8 for still version.
Fig. 21

Fig. 21. Spatiotemporal interpolation results for eastern red bats along a forest fragment in southeast Nebraska. Maps were made by kriging with a normal score transformation with bins containing the call count groupings in half-hour increments spanning a period of four nights. Refer to Fig. 8 for still version.
Fig. 22

Fig. 22. Line plot with error bars of insect volumetric measurements from light traps at increasing distances from tree cover on 5 agricultural sites (red) and 1 tallgrass prairie (blue) in southeast Nebraska. Sites were sampled once over a 5 hour period beginning at sunset. Traps were placed at 20m, 220m, and 420m from crop field edges, and contents from each trap measured in a 250mL graduated cylinder.