|
|
|
Copyright © 2004 by the author(s). Published here under license by The Resilience Alliance.
Go to the pdf version of this article.
The following is the established format for referencing this article:
Wear, D., J. Pye, and K. Riitters. 2004. Defining conservation priorities using fragmentation forecasts. Ecology and Society 9(5): 4. [online] URL: http://www.ecologyandsociety.org/vol9/iss5/art4/
Report, part of Special Feature on Urban Sprawl Defining Conservation Priorities Using Fragmentation Forecasts David Wear, John Pye, and Kurt Riitters
U.S. Forest Service
- Abstract
- Introduction
- Study Area
- Methods
- Data
- Results
- Discussion and Speculation
- Responses to this Article
- Literature Cited
Methods are developed for forecasting the effects of population and economic growth on the distribution of interior forest habitat. An application to the southeastern United States shows that models provide significant explanatory power with regard to the observed distribution of interior forest. Estimates for economic and biophysical variables are significant and consistent with theory. Forecasts of interior forest based on the population and economic growth projected for the region are displayed by ecological section and province and by metropolitan statistical area (MSA). Loss of interior forests is expected to be especially high in certain ecological sections, including the southern Appalachian Piedmont in North and South Carolina, the Gulf prairies and marshes in Texas, and the Florida coastal lowlands. Sixty-six percent of loss of interior forests will be in urban counties, which highlights the conservation importance of the urbanizing fringe of several cities. Among the ten MSAs that will lose the most interior forest, seven are found in Florida. These forecasts provide a mechanism for assigning priorities and targeting areas for more detailed study and for conservation efforts.
KEY WORDS: fragmentation, interior forest, land use forecasts, North Carolina, South Carolina, Florida, Texas.
Published: September 14, 2004
Conservation concerns have long focused not only on the availability of habitat but also on the structural integrity of that habitat. One index of structural integrity is habitat contiguity, which can be measured in terms of the relative isolation of forest patches, i.e., an island biogeography concept, or the amount of interior forest cover that is functionally distinct from edge forest, i.e., a landscape ecology concept. Effective conservation calls for targeting actions not only toward areas in which habitat conditions are currently limiting but also toward those in which conditions are expected to become limiting in the future. This paper addresses the need to look ahead when defining conservation priorities by developing forecasting methods that identify the areas in which population and economic growth may give rise to structural changes in forest cover.
Our research focuses on the southeastern United States, where land is largely in private ownership (89% according to Conner and Hartsell 2002) and where land and resource use has long been dynamic and organized by the individual decisions of hundreds of thousands of private landowners. A recent interdisciplinary assessment of forest conditions in this region (Wear and Greis 2002) indicates that, although expanding timber management will have ongoing effects on forest composition, anticipated urbanization coupled with low-density residential development could have the most substantial effects on the structure and function of forest ecosystems over the next 40 yr. Wear and Greis (2002) project that about 12 x 106 acres of forest or about 6% will be converted to an urban use over this time span. Most of these changes will be focused in a relatively small portion of the southeast, including the southern Appalachian Piedmont and the Atlantic coastal flatwoods.
Changes in land use alone may not capture the ecological implications of human development in the region. For example, in areas that are currently heavily forested, relatively large changes in forest area may have little effect on the structural qualities of the forest, i.e., on the share of forest that is interior vs. edge, whereas, in areas with less forest, relatively small changes in forest area could give rise to significant structural changes. The objective of the research presented here is to develop methods for forecasting changes in forest structure related to several driving factors, including population and economic activity, and conditioned on the biophysical characteristics of an area. The modeling and forecasting are conducted at fairly broad scales ranging from counties up to ecological provinces, and provide a tool for assigning priorities to conservation concerns and efforts at those scales.
Our approach builds on methods used to forecast land use proportions at the county level (e.g., Hardie et al. 2000). We use cross-sectional observations of land-cover configurations summarized at the county level to test hypotheses about the effects of driving variables on existing land-cover configurations. The resulting multiple-equation models, estimated using regression techniques, provide the basis for examining potential future impacts on forest configuration. These are measured by first gauging the current sensitivity of the forest structure to marginal changes in the human population and then forecasting changes in forest configuration based on demographic and economic projections.
For this study, we focus on a single measure of forest configuration: the proportion of land that is considered "interior forest cover." Interior forest is defined as a forest cell that is surrounded by other forest cells. It is therefore inversely related to the degree of fragmentation of a landscape and provides a straightforward summary of forest-cover conditions on a pixilated land-cover map. Although other measures of fragmentation are available, interior forest area provides a readily constructed and useful summary measure for analysis at such a broad scale (Riitters et al. 2000, 2002).
It is implicit in our research approach that the amount of interior forest serves as an indicator of available habitat for certain fragmentation-sensitive species. Species respond to forest fragmentation in different ways and to different degrees (Andrén 1994, Debinski and Holt 2000, Bissonette and Storch 2002). The amount of suitable habitat in a given area depends not only on the amount but also on the arrangement of forest in relation to the life-history requirements of the species who occupy it (Riitters et al. 1997). Maintaining total forest-dependent biodiversity requires having suitable fragmented (edge) habitats as well as unfragmented (interior) habitats. Regional assessments often focus on interior forest because it is a sensitive indicator of fragmentation (e.g., Heilman et al. 2002, Heinz Center 2002) and because edge habitat is often widely available (e.g., Riitters et al. 2002). As development proceeds, edge habitat becomes more plentiful and interior habitat becomes more scarce. Our approach could be extended to evaluate the amounts and locations of edge habitat created by urbanization and road-building, or to help evaluate the future impacts on a wide range of ecological processes that are affected by fragmentation (Saunders et al. 1991).
Our study area includes 12 states in the southeastern United States (Fig. 1) from Texas and Arkansas eastward to the Atlantic Coast and bounded on the north by Kentucky and Virginia. We include only the forested portion of Texas in our analysis and exclude the Everglades subregion of Florida. The National Hierarchical Framework for Ecological Units (ECOMAP 1993) indicates that this region encompasses 32 ecological sections and 10 ecological provinces. This ecological diversity is coupled with broad social and economic diversity ranging from remote, relatively isolated communities with rural economies to large metropolitan areas.
The southeastern United States has experienced a long history of land use and economic change, with much of the land switching from forest to agriculture and back to forest over the past 200 yr (Wear 2002). Today, forest covers more than 60% of most states in the region, and 89% of all forest land is in private ownership (Conner and Hartsell 2002). Ongoing changes continue to reshape the landscape, with economic growth driving urbanization; five of the region's states are among the top 10 in current rates of urbanization (NRI 2002). This region of the United States also produces more timber products than any other entire country in the world. As a result, roughly 16% of the region's forests are managed in an intensive manner, and this type of management is expected to expand in the future. Wear and Greis (2002) outline these and several other forces of change in the region.
Our general approach is to model the observed proportion of interior forest within a county as a function of a set of independent variables. Relationships are estimated using regression techniques that allow for hypothesis testing using t and F statistics. The fitted equations are then used to forecast where and how interior forest may change in the future.
We estimate models at the county level because this is the finest scale at which all the explanatory variables are recorded. After we construct forecasts at the county level, we then aggregate the results for broader subdivisions of the region. In particular, we construct estimates for ecological sections and provinces and for metropolitan statistical areas (MSAs). This allows us to identify the subregions of the south that are most likely to experience a change in forest cover in the future.
The proportion of a county classified as interior forest (s) can be modeled as a simple function of the proportion of the county that is forested (f) and the distribution of nonforest uses within the county, i.e., the degree to which the county is fragmented. We approximate this fragmentation using the proportion of the county's pixels that contain a road (R). These variables are determined by a set of independent variables that relate to human occupancy and land uses. Assuming that a logistic cumulative distribution function applies to the probability that a pixel is interior forest, we can apply the aggregate or grouped logit model to the proportion of interior forest:
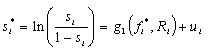 | (1) |
where i indexes the county, si* is the log-odds measure of the proportion of interior forest cover, fi* is the log-odds measure of the proportion of total forest cover, R is a measure of road density, and ui is an error term. We anticipate that interior forest is positively related to forest cover and negatively related to road density. The log-odds transformation gives rise to heteroscedasticity that can be addressed using weighted least squares where the variance of ui is 1/(Aisi[1-si]) and Ai is the area of the county (see Madalla 1983).
The two explanatory variables in Eq. 1, forest cover and road density, are determined by a number of socioeconomic and biophysical factors. In particular, the area of forest is influenced by the variables that affect the allocation of land to forest or nonforest uses.
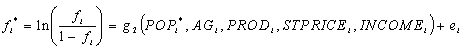 | (2) |
where POP is the population density of the county, AG is the market value per acre of agricultural products sold, PROD is the proportion of land use in Land Capability Classes I and II, STPRICE is the price of sawtimber, and INCOME is the average household income. POP and INCOME capture factors that shift land to urban uses, whereas AG, PROD, and STPRICE reflect forces that allocate rural land between agricultural and forest uses. Equation 2 is also subject to heteroscedastic errors with variance of ei equal to 1/(Aifi[1-fi]), which can be corrected by using weighted least squares.
Road density is modeled as follows:
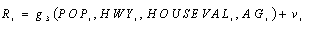 | (3) |
where HWY is a dummy variable that indicates whether or not the county contains a U.S. highway, and HOUSEVAL is the average value of houses in the county. We posit that all of these variables are positively related to road density.
Equations 1, 2, and 3 can be viewed as a system with simultaneous equation bias arising from endogenous variables on the right-hand side of Eq. 1. Their respective error terms (u, e, and v) are also likely to be correlated, so that coefficient estimates of individual equations would be inefficient. To address these concerns, we estimate these equations as a system using three-stage least squares and the weighting scheme identified above.
We also expect that these relationships may be influenced by biophysical and topographic conditions that differ between ecological provinces. Accordingly, we include a series of dummy variables in the equations that indicate the province within which the county is located. We combine mountain provinces and exclude counties in the Everglades province. Dummy variables were constructed for (1) southeastern mixed forest; (2) outer coastal plain; (3) lower Mississippi riverine; (4) mountain broadleaf, including Appalachian, Ouachita, and Ozark sections; and (5) prairie parkland provinces. The null case, i.e., all dummies equal to zero, is the eastern broadleaf forest province. These dummy variables are included on the right-hand side of all three equations.
Our objective is to evaluate the effect of various driving factors on the proportion of a county that is interior forest. To provide a scale-free evaluation of the sensitivity of one variable to change in another, we use coefficient estimates to calculate elasticities. Elasticities are commonly used in economics and are defined as the ratio of the proportional change in the dependent variable, e.g., interior forest, and the associated proportional change in the independent variable, e.g., population. These measures provide a useful means of comparing sensitivity across dissimilar units such as counties of different sizes.
We start by examining the estimated relationship between dependent and independent variables. Because of the nonlinear transformations used to construct Eqs. 1 and 2, this requires information from all three equations. By the chain rule of differential calculus, the effect of a marginal change in population on the proportion of interior forest is defined as:
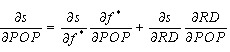 | (4) |
Note that the effect on the proportion of interior forest needs to be derived from the effects on the log-odds measure in Eq. 1:
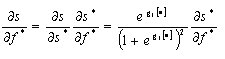 | (5) |
To compare the sensitivity of counties to a marginal change in population, we calculate the elasticity of forest interior with respect to the driving variable, that is, the proportional change in interior forest with respect to a proportional change in population.
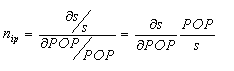 | (6) |
Elasticities are also calculated for the effects of changes in forest area and road density on interior forest. We calculate these for each county but also at the point of means, i.e., with all the independent variables set to their mean values, to provide a general measure of responsiveness.
Forest land area and fragmentation were measured using the National Land Cover Data (NLCD) land-cover map derived from 1992 Landsat Thematic Mapper data (Vogelmann et al. 2001). Four NLCD forest classes, i.e., coniferous forest, deciduous forest, mixed forest, and woody wetlands, were combined into one "forest" class for this analysis. The spatial resolution of the NLCD map is 0.09 ha/pixel, and the accuracy per pixel of the forest vs. nonforest classification is approximately 90% for the study area. (Yang et al. 2001). The proportion of each county that was forested was estimated by the proportion of pixels in a county that were classified as forest. These area estimates may differ from official area statistics because of differences in the definition of forest, sampling procedures, mapping conventions, and year of observation.
The fragmentation analysis used the spatial methods described by Riitters et al. (2002). Briefly, the NLCD map was reclassed as (1) forest by the method described above, (2) nonforest, and (3) missing, e.g., water, beach; the missing class was subsequently ignored in all calculations. Each 0.09-ha pixel was then characterized in terms of the connectivity of all forest contained within the 7.29-ha2 window surrounding that pixel. Connectivity was estimated by the Pff statistic (Riitters et al. 2000) defined as the number of "pixel edges" in a window that separate two forest pixels, i.e., forest-forest "pixel edges," divided by the total number of "pixel edges" that have a forest pixel on at least one side, i.e., forest-forest "pixel edges" plus forest-nonforest "pixel edges." Roughly, Pff is a measure of the conditional probability that a pixel next to a forest pixel is also forest. The range of Pff is [0, 100], and larger values indicate less fragmentation. The statistic was set to zero for windows containing no forest. County-level statistics were obtained by constructing the frequency distributions of the Pff statistic for each county and calculating the proportion of pixels within each county that exceeded a specified threshold Pff value. We refer to those pixels as "interior" forest, and we varied the threshold value from 70 to 100% to test sensitivity to this aspect of the fragmentation model.
When the threshold is set at 100%, an interior forest pixel is surrounded by a 9 pixel x 9 pixel, i.e., 7.29-ha, landscape that contains only forest pixels. In that case, the interior pixel is at least 120 m, i.e., 4 pixels, away from the nearest forest edge. Forest pixels that do not meet a threshold value of 100% have at least some forest edge closer than 120 m. The distance over which forest edges have an effect varies with circumstances and the ecological process in question (Gehlhausen et al. 2000). Different edge effect distances suggest the use of landscape windows of different sizes, and, generally speaking, one finds less interior forest and more "edge-affected forest" when using landscape windows larger than 7.29 ha (Riitters et al. 2000, 2002). In other words, measures of fragmentation are scale-dependent. Our choice of a 7.29-ha landscape scale was based on the approximate scale at which humans decide land uses and not on the specific life-history requirement, e.g., home range size, of any particular species (McIntyre and Hobbs 1999).
Road density was measured using a 1995 modification of the Bureau of the Census TIGER/Line files that identified public roads ranging from interstate highways to trails for vehicles with four-wheel drive (Geographic Data Technology 2002). Although the database includes remarkable detail for small roads in rural areas, it does not include all the roads that are used to access the forest for harvest, fire protection, right-of-way management, and other similar activities. To facilitate comparisons with the land-cover and fragmentation maps, the road map was converted to a grid of road and nonroad pixels at a spatial resolution of 0.09 ha/pixel and summarized for each county (Riitters and Wickham 2003). No distinctions were drawn between the type of road, traffic volume, or other factors. Road density was thus measured in terms of the proportion of total area that contained at least one road segment. In comparison to expressing road density in terms of kilometers of road per square kilometer of land, this approach gives less weight to heavily roaded urban areas within counties and, as a result, is more sensitive to the degree to which roads tend to pervade rural areas in which forest fragmentation is more likely to be located.
Other dataIndependent variables for Eqs. 1, 2, and 3 are defined as follows:
- PROD, our measure of potential productivity, is the proportion of the county that is in Land Capability Classes I and II as defined by the Natural Resource Conservation Service. Land Capability Classes are categories of soil groupings defined by their ability to produce cultivated crops and pasture plants. Soils in classes I and II constitute those that do not have severe limitations restricting the choice of plants (U.S. Department of Agriculture 2003).
- STPRICE, stumpage price, is the price of standing pine sawtimber in dollars per thousand board feet in 1992. These data are taken from the price reporting service, Timber Mart South, and are reported for two or three multiple-county regions in each state in our study area.
- AG is the revenue per acre from all agricultural activities reported by county in the 1992 Census of Agriculture.
- INCOME is the median household income for the county from the U.S. Department of Commerce, Bureau of Economic Analysis.
- HOUSEVAL is the median house value for the county in 1992 as reported by the U.S. Department of Commerce, Bureau of the Census.
- HWY is a dummy variable that is equal to one if a U.S. highway is present in the county; otherwise its value is zero. This variable was developed using an algorithm in ARCINFO (ESRI).
- POP is the population density of the county based on estimates from the U.S. Department of Commerce, Census Bureau, for 1992.
Estimates of the three-equation system (Table 1) show a high degree of fit and several significant explanatory variables (we evaluate significance at the P = 0.05 level). The equation for interior forest has an adjusted R2 of 0.83, and both the forest cover and the roads variables are significant. Consistent with expectations, interior forest cover is positively related to the forest cover and is negatively related to the road density in the county. The relationship between interior forest and these two explanatory variables is not significantly different in the six ecological provinces, i.e., all five ecoregion dummy variables are not significantly different from zero.
The adjusted R2 for the road density equation is 0.68. Road density within a county is significantly and positively related to the presence of a U.S. highway, population density, and the average value of housing. The value of agricultural products does not have a significant effect on road density. Counties in the mountain broadleaf ecological provinces have higher road densities and outer coastal plain and lower Mississippi provinces have lower road densities than do comparable counties in the eastern broadleaf forest province (the null case). The relationships between explanatory variables and road density are not significantly different for the southeastern mixed forest and prairie parkland provinces.
The proportion of forest cover is negatively related to population density and household income, which are indicators of urbanization, and negatively related to the value of agricultural products and site productivity, which are indicators of agricultural uses of rural land. Timber prices do not have a significant effect on forest cover. The counties in the outer coastal plain and the lower Mississippi provinces have less forest cover than do similar counties in the eastern broadleaf forest province. Counties in the mountain broadleaf province have more forest cover than for the null case. The adjusted R2 for the forest-cover equation is 0.34.
We also estimated models for alternative definitions of interior forest: Pff > 90, Pff > 80, and Pff > 70. As the definition becomes less restrictive, i.e., moves toward Pff > 70, the explanatory power of the model increases, and the adjusted R2 for the interior forest equation increases from 0.83 to 0.99. The roads variable in the interior forest-cover equation is not significantly different from zero for the Pff > 80 and Pff > 70 models, indicating that forest cover alone explains most of the variation in interior forest for these cases.
ElasticitiesWe summarize the effects of independent variables on the amount of interior forest using elasticities as defined by Eqs. 4–6. We calculated elasticities of interior forest with respect to road density and forest cover. We also calculated the elasticity of interior forest with respect to population, which involves combining the effects of population on road density and forest cover with these other two elasticities. We calculated these elasticities at the point of means, i.e., with all variables assigned their mean values, for the explanatory variables in our data set.
As expected, the elasticity of interior forests with respect to forest cover is strongly positive (Table 2). The elasticity of 0.66 indicates that a 1% increase (decrease) in forest would, with other variables held constant, give rise to a 0.66% increase (decrease) in interior forest. The elasticity of interior forest with respect to road density is -0.36, indicating that a 1% increase in road density would reduce interior forest cover by 0.36%. The elasticity of interior forest with respect to population density (ηip) is equal to -0.14 at the point of means.
We also calculated the ηip for each observation in our data set and then plotted these against population density (Fig. 2). ηip is near zero for very low values of population. As population increases, so does the magnitude ηip. The six most populous counties of the region have ηip values of between -2 and -3. Roughly 90% of counties have elasticities between -0.00 and -0.5.
We decomposed the ηip to examine the relative effects that population change would have on interior forest through (1) decreased forest cover and (2) increased road density. We calculated total and decomposed elasticities at the point of means, then varied population density over the range shown in Fig. 3. The forest cover effect dominates the road density effect across the entire range of population densities (Fig. 3). The forest cover component of ηip is 65% greater than the road density component.
We also calculated elasticities for alternative definitions of interior forest (Table 2). As the definition becomes less restrictive, i.e., moves from Pff = 100 to Pff > 70, the effect of population density on interior forest is greatly reduced. At Pff = 100, ηip is -0.14. At Pff > 70, ηip is -0.03.
The relationship between population density and ηip for Pff = 100 is also seen in its spatial distribution (Fig. 4). The counties in the vicinity of Atlanta (Georgia), Houston (Texas), and Louisville (Kentucky) have the highest elasticities. A large contiguous area of elevated elasticities is found in the North Carolina portion of the southern Appalachian Piedmont. This area is anchored at the eastern extreme by the city of Raleigh, North Carolina. Charlotte anchors the western end. A similarly sized contiguous area of elevated elasticities is found in central Florida, in the counties around Tampa Bay and Orlando. Also apparent are large portions of the south in which interior forest is relatively insensitive to marginal changes in population. This includes the upper coastal plain encompassing Georgia and Alabama and most of the state of Mississippi. The entire state of Arkansas shows relatively low elasticities.
Forecasts
We forecast changes in interior forest for each county in the study area based on population density forecasts to the year 2020; it was assumed that relative returns to agricultural and timber production would remain at current levels. Under this scenario, the south as a whole is forecast to lose 747,000 ha, or about 2.12%, of interior forest cover (Table 3).
These changes are not constant across the region, consistent with the spatial variation in the estimated elasticities. Among ecological sections, the southern Appalachian Piedmont would lose the most area of interior forest cover at 173,166 ha. The Gulf prairies and marshes in Texas have a very small area of interior forest and represent the ecological section with the greatest loss of interior forest at 56.7% (Table 3). The second and fourth greatest reductions are found in the eastern and western Florida coastal lowlands, respectively. Aggregation to the ecological province level indicates that the outer coastal plain would experience the greatest reduction in interior forest (Table 4). All ecological sections with losses forecast at greater than 2% were located either along the Atlantic and Gulf coastal plain or in the upland areas of North Carolina, Virginia, Tennessee, and Kentucky.
The spatial distribution of forecast changes in interior forest indicates that losses will be focused in distinct subregions (Fig. 5). Similar to the elasticity results, losses of interior forest are forecast to be concentrated in northern Georgia, central North Carolina, and much of peninsular Florida. Large areas of interior forest loss are also forecast for counties around Nashville and Knoxville, Tennessee, and counties along the Gulf of Mexico from the panhandle of Florida to Louisiana. These areas of forecast loss are focused on urbanizing areas and, without exception, are organized along interstate highway corridors.
We further examine the distribution of forecast losses of forest interior between urban and rural counties. Urban counties were defined on June 23, 1998, as those attached to metropolitan statistical areas (MSAs) by the Office of Management and Budget, and rural counties were defined as the remainder. The MSA counties contain 492,690 ha or 66% of the total forecast loss of 747,744 ha. Heavily impacted MSAs are concentrated in Florida (Table 5). The Tampa-St. Petersburg-Clearwater MSA is forecast to lose 34.5% of its interior forest, and seven of the 10 MSAs with the highest percentage losses are found in Florida. Columbia (South Carolina), Atlanta (Georgia), and Raleigh-Durham-Chapel Hill (North Carolina) round out the top 10.
The model presented here provides a mechanism for interpreting the effects of potential growth and development on ecologically meaningful outcomes. It allows analysts to identify where and how interior forest cover may change over time and, in particular, to identify where it may become increasingly scarce in the near future. This type of approach could prove useful for targeting areas for conservation efforts. We speculate on a four-step approach for broad-scale targeting of conservation activities in areas like the U.S. south, where most lands are privately held and ecological conditions are strongly shaped by economic processes.
Reserves are typically viewed as the preferred mechanism for protecting biodiversity, but reserves alone are not enough to protect biodiversity because they have generally not been designed to provide a representative sample of biota (Margules and Pressey 2000). As a result, future protection of biodiversity and other ecological attributes will need to focus on the management and condition of private land in many parts of the world (Margules and Pressey 2000, Theobold 2003). Indeed, in the U.S. south, only 11% of forests are in pubic ownership, and many of these areas are managed for timber production (Wear and Greis 2002). It is unlikely that the current configuration of reserves is organized in a way that supports long-term conservation goals, although an exception may be the southern Appalachian highlands, where about 25% of land is in public ownership (U.S. Forest Service 1996). Across the south, traditional conservation activities such as tree planting and stewardship planning have been largely driven by financial inducements for private landowners and provided without the spatial targeting needed to address biodiversity concerns.
In the U.S. south, conservation activities are increasingly organized by the private sector, with nongovernment organizations either purchasing lands outright or acquiring property rights for ecologically significant aspects of land through conservation easements. In contrast to regions in which a central planning/governmental approach to conservation activities can be applied (see, for example, Pressey et al. 2000), conservation activities are accomplished through a decentralized approach involving a variety of actors. Within such an institutional structure, information that allows these actors to identify the critical ecological assets most threatened by ongoing development is extremely valuable.
Our methods define one way of tracking ecological structures that are changing in response to human population growth and economic developments at broad scales across the region. The identification of specific conservation targets within these broad areas would require additional work at a finer scale (e.g., Tankersley and Orvis 2003). Nevertheless, the methods described here could provide a mechanism for defining conservation priorities for the region at a broad scale. The forecasts of interior forest change can be viewed as a risk measure indicating where human activities are more likely to change ecosystem structure, similar to the risk indicators developed at a much finer scale by Theobold (2003). This provides a first step in setting priorities: defining where the structure is likely to change and where it is likely to be relatively stable. Table 3 identifies four ecological sections of the south where 5% or more of existing interior forest is forecast to be lost by 2020, but it also identifies 16 ecological sections, or about half of the sections in the southeastern United States, where less than 1.5% would be lost. Areas that are found to be essentially stable could be excluded from further detailed analysis, allowing analysts to focus their efforts on that portion of the landscape that is more likely to change without some intervention.
A second step in defining conservation priorities involves examining the ecological condition of the broad areas with relatively high threat levels. Indicators such as numbers of imperiled species highlight where ongoing change may have the most impact on biodiversity. Although a detailed assessment of ecological scarcity is beyond the scope of this paper, we use the recently completed Southern Forest Resource Assessment (Wear and Greis 2002) to illustrate the approach. We highlight four areas with high levels of imperiled species and other ecological concerns.
- The Gulf prairies and marshes province of eastern Texas is forecast to lose more than half of its interior forest. This is one of the smallest ecological sections in the study area and one with low forest cover. Areas within this
province contain a large number of imperiled plant species (Owen 2002). Because of its close proximity to the Houston metropolitan area, concerns focus on a relatively small number of forest tracts threatened by urbanization.
- The eastern and western Florida coastal lowlands and coastal plain and flatwoods represents a much larger area that is both heavily forested and experiencing fragmentation. Forecast losses of interior forests reflect urbanization pressure in rapidly developing coastal areas. Although these areas are highly desirable because of amenities and recreational opportunities, they also have high ecological value. The Southern Forest Resource Assessment (SFRA) finds the highest concentration of imperiled amphibians, reptiles, and birds in these coastal regions (Griep 2002). The SFRA also identifies this as one of three hotspots for endangered plant species in the south (Owen 2002).
- The southern Appalachian Piedmont is forecast to lose a large portion of its interior forest by 2020. Because this section had relatively low amounts of interior forest to start with, the implications of forecast losses may be substantial. Within this region, change is focused in the greater Atlanta area and through much of central North Carolina. The SFRA emphasizes concerns regarding habitat for neotropical migrant birds in the Piedmont (Griep 2002), and our findings indicate that the availability of interior habitat may become more limiting in the future.
- Several ecological sections in Tennessee including the central ridge and valley, southern Cumberland Plateau, and southern ridge and valley are forecast to lose relatively high proportions of interior forest, making this state another one with concerns about fragmentation. Imperiled endemic species, especially on the southern Cumberland Plateau, may be adversely affected by these changes (Griep 2002).
The third step in assigning priorities for conservation is to define the subareas within these threatened ecological sections that are changing most rapidly. In general, our findings indicate that concerns about forest fragmentation are focused on urban areas. Elasticities of interior forest with respect to population increase with the population density of the area. Metropolitan statistical areas (MSAs) are forecast to sustain 66% of the forest interior losses in the U.S. south, suggesting that conservation priorities may need to be assigned to the urbanizing fringes of the MSAs within threatened ecological sections.
The fourth step in setting conservation priorities is a careful cost:benefit analysis of alternative conservation strategies. In urbanizing areas, the impacts of interior losses, and therefore the potential returns on conservation investments, are likely to be high, but the value of land for development, and therefore the cost of conservation, is also quite high. The gradient of land prices moving from the edge of development outward typically defines a tradeoff between the location of activities and the quantity of land that can be protected. A comparison of this cost:quantity tradeoff with the potential effectiveness of protection as a function of the size of holdings could then be used to identify the optimal location for land purchases and for conservation easements. In urbanizing areas, high costs and rates of irreversible change imply that tradeoffs between conservation costs and benefits may be quite large and spatially variable. Accordingly, substantial benefits may accrue to focused and intensive cost:benefit assessments of conservation alternatives for this portion of the landscape, i.e., areas around rapidly developing MSAs.
Future research could be focused in a number of areas. Other measures of forest structure, including multiple attribute measures, could be evaluated and might prove more useful as indices of ecological change. The links between forest cover measures and habitat conditions in specific places, including a definition of critical thresholds of damage, could enhance the usefulness of these methods. A framework for effective cost:benefit analysis as described in step four above needs to be developed. As with any forecasting effort, empirical validation of forecasts is highly desirable. This will become feasible with the completion of a second National Land Cover Data mapping project in the next few years.
Responses to this article are invited. If accepted for publication, your response will be hyperlinked to the article. To submit a comment, follow this link. To read comments already accepted, follow this link.
Andrén, E. 1994. Effects of habitat fragmentation on birds and mammals in landscapes with different proportions of suitable habitat: a review. Oikos 71:355-366.
Bissonette, J. A., and I. Storch. 2002. Fragmentation: is the message clear? Conservation Ecology 6(2):14. [online] URL: http://www.consecol.org/vol6/iss2/art14.
Conner, R. C., and A. J. Hartsell. 2002. Forest area and conditions. Pages 357-402 in D. N. Wear and J. G. Greis, editors. The southern forest resource assessment: technical report. U.S. Forest Service General Technical Report SRS-53.
Debinski, D. M., and R. D. Holt. 2000. A survey and overview of habitat fragmentation experiments. Conservation Biology 14:342-355.
ECOMAP. 1993. National hierarchical framework of ecological units. U.S. Forest Service, Washington, D.C., USA.
Gehlhausen, S. M., M. W. Schwartz, and C. K. Augspurger. 2000. Vegetation and microclimatic edge effects in two mixed-mesophytic forest fragments. Plant Ecology 147:21-35.
Geographic Data Technology. 2002. Dynamap/2000 user manual. Geographic Data Technology, Lebanon, New Hampshire, USA.
Griep, M. T. 2002. Maintaining species in the South. Pages 113-150 in D. N. Wear and J. G. Greis, editors. The southern forest resource assessment: technical report. U.S. Forest Service General Technical Report SRS-53.
Hardie, I., P. Parks, P. Gottlieb, and D. Wear. 2000. Responsiveness of rural and urban land uses to land rent determinants in the south. Land Economics 76(4):659-673.
Heilman, G. E., Jr., J. R. Strittholt, N. C. Slosser, and D. A. Dellasala. 2002. Forest fragmentation of the conterminous United States: assessing forest intactness through road density and spatial characteristics. BioScience 52:411-422.
Heinz Center (The H. John Heinz III Center for Science, Economics and the Environment). 2002. The state of the nation's ecosystems: measuring the lands, waters, and living resources of the United States. Cambridge University Press, Cambridge, UK.
Judge, G. G., W. E. Griffiths, R. C. Hill, W. E. Lutkepolh, and T. C. Lee. 1985. The theory and practice of econometrics. Wiley, New York, New York, USA.
Maddala, G. S. 1983. Limited-dependent and qualitative variables in econometrics. Cambridge University Press, New York, New York, USA.
Margules, C. R., and R. L. Pressey. 2000. Systematic conservation planning. Nature 405:243-253.
McIntyre, S. and R. Hobbs. 1999. A framework for conceptualizing human effects on landscapes and its relevance to management and research models. Conservation Biology 13:1282-1292.
Owen, W. 2002 The history of native plant communities in the south. Pages 47-62 in D. N. Wear and J. G. Greis, editors. The southern forest resource assessment: technical report. U.S. Forest Service General Technical Report SRS-53.
Pressey, R. L., T. C. Hager, K. M. Ryan, J. Schwarz, S. Wall, S. Ferrier, and P. M. Creaser. 2000. Using abiotic data for conservation assessments over extensive regions: quantitative methods applied across New South Wales, Australia. Biological Conservation 96:55-82.
National Cartography and Geospatial Center. 1998. Instructions for collecting the 1997 National Resources Inventory data. Available online at www.ncg.nrcs.usda.gov/nri/inst_archive.php.
Riitters, K. H., R. V. O'Neill, and K. B. Jones. 1997. Assessing habitat suitability at multiple scales: a landscape-level approach. Biological Conservation 81:191-202.
Riitters K. H., J. D. Wickham, R. V. O'Neill, K. B. Jones, and E. R. Smith. 2000. Global-scale patterns of forest fragmentation. Conservation Ecology 4(2):3. [online] URL: http://www.consecol.org/Journal/vol4/iss2/art3/.
Riitters, K. H., J. D. Wickham, R. V. O'Neill, K. B. Jones, K.B., Smith, J. W. Coulston, T. G. Wade, and J. H. Smith. 2002. Fragmentation of continental United States forests. Ecosystems 5:815-822.
Riitters, K. H., and J. D. Wickham. 2003. How far to the nearest road? Frontiers in Ecology and the Environment 1:125-129.
Rudis, V. A. 1999. Ecological subregion codes by county, coterminous United States. U.S. Forest Service General Technical Report GTR-SRS.
Saunders, D. A., R. J. Hobbs, and C.R. Margules. 1991. Biological consequences of ecosystem fragmentation: a review. Conservation Biology 5:18-32.
Tankersley Jr., R., and K. Orvis. 2003. Modeling the geography of migratory pathways and stopover habitats for neotropical migratory birds. Conservation Ecology 7(1):7. [online] URL: http://www.consecol.org/vol7/iss1/art7.
Theobald, D. M. 2003. Targeting conservation action through assessment of protection and exurban threats. Conservation Biology 17(6):1624-1637.
U.S. Department of Agriculture. 2003. National soil survey handbook, title 430-VI. Available online at: http://soils.usda.gov/technical/handbook/.
U.S. Forest Service. 1996. The southern Appalachian assessment social/ciultural/economic technical report. U.S. Forest Service, Southern Region, Atlanta, Georgia, USA.
Vogelmann, J. E., S. M. Howard, L. Yang, C. R. Larson, B. K. Wylie, and N. Van Driel. 2001. Completion of the 1990s national land cover data set for the conterminous United States from Landsat Thematic Mapper data and ancillary data sources. Photogrammetric Engineering and Remote Sensing 67:650-662.
Wear, D. N. 2002. Land use. Pages 153-187 in D. N. Wear and J. G. Greis, editors. The southern forest resource assessment: technical report. U.S. Forest Service General Technical Report SRS-53.
Wear, D. N., and P. Bolstad. 1998. Land use changes in southern Appalachian landscapes: spatial analysis and forecast evaluation. Ecosystems 1(6):575-594.
Wear, D. N., and J. G. Greis. 2002. The southern rorest resource assessment: summary report. U.S. Forest Service General Technical Report SRS-54.
Yang, L., S. V. Stehman, J. H. Smith, and J. D. Wickham. 2001. Thematic accuracy of MRLC land cover for the eastern United States. Remote Sensing of the Environment 76:418-422.
Address of Correspondent:
David Wear
U.S. Forest Service
3041 Cornwallis Road
Research Triangle Park, North Carolina 27709 USA
Phone: (919) 549-4011
Fax: (919) 549-4047
dwear@fs.fed.us
![]()