|
|
|
Copyright © 2004 by the author(s). Published here under license by The Resilience Alliance.
Go to the pdf version of this article.
The following is the established format for referencing this article:
Shine, R., M. Lemaster, M. Wall, T. Langkilde, and R. Mason. 2004. Why did the snake cross the road? Effects of roads on movement and location of mates by garter snakes (Thamnophis sirtalis parietalis). Ecology and Society 9(1): 9. [online] URL: http://www.ecologyandsociety.org/vol9/iss1/art9/
A version of this article in which text, figures, tables, and appendices are separate files may be found by following this link.
Report Why Did the Snake Cross the Road? Effects of Roads on Movement and Location of Mates by Garter Snakes (Thamnophis sirtalis parietalis) Richard Shine1, Michael Lemaster, Michael Wall, Tracy Langkilde, and Robert Mason
1University of Sydney
- Abstract
- Introduction
- Methods
- Results
- Discussion
- Responses to this Article
- Acknowledgments
- Literature Cited
If animals avoid road surfaces or are unable to follow conspecific trails across such surfaces, previously continuous populations may be fragmented. We gathered data on the effects of a small (4-m wide) gravel road on the behavior and trail-following abilities of garter snakes (Thamnophis sirtalis parietalis) in Manitoba, central Canada. As expected, the road surface had less vegetation cover, a more open canopy and, thus, higher incident radiation than did the surrounding grassland. Contrary to expectations, however, substrate temperatures were lower on the road than in its surrounds, because of the higher reflectivity of the road's surface. On a nearby asphalt road, substrate temperatures were relatively high on the road surface only in the evening, as surrounding areas cooled. Focal sampling showed that snakes avoided the gravel road, typically changing direction when they encountered it. If they crossed the road, they did so by the shortest possible route (straight across). Mate-searching male snakes were less able to follow substrate-deposited pheromonal trails left by females if those trails crossed a road than if the trails were entirely within the surrounding grassland. Thus, roads may significantly modify snake movement patterns, as well as the ability of males to locate reproductive females. Our study provides the first detailed information on the effects of roads on snake behavior.
KEY WORDS: behavior, connectivity, habitat fragmentation, pheromones, reproduction, reptile.
Published: March 17, 2004
Human activities have modified natural habitats in many ways. The most dramatic changes involve widespread degradation of entire areas, such as a shift from forest to agricultural use. However, a more pervasive influence is the construction of linear open areas—tracks, paths, and roads—through previously continuous habitat (Forman 2000). These open areas may provide ecological situations that differ profoundly from those of the surrounding habitat; for example, sun penetration may modify floral composition, nest-site availability, and the like (Ries et al. 2001, Shine et al. 2002, Riitters and Wickham 2003). Predictably, animals exhibit a wide range of behavioral responses to the novel environments provided by roads: some taxa avoid open areas, whereas others use these paths as invasion routes (May and Norton 1996, Seabrook and Dettmann 1996, Stiles and Jones 1998). Roads that constitute behavioral barriers to animal movement may effectively fragment populations in otherwise continuous habitat (Mader 1984, Reh 1989, Fehlberg 1994). All these impacts may influence the continued viability of populations and, thus, have attracted increasing study from conservation biologists (Mader 1984, Reh 1989, Alexander and Waters 2000, Jones 2000, Lodé 2000).
Reptiles have attracted less scientific attention in this respect than have groups such as ungulates (e.g., Waring et al. 1991, Groot Bruinderink and Hazebroek 1996, Putman 1997, Clevenger et al. 2001, Nielsen et al. 2003, Rea 2003). Most studies of road effects on reptiles have focused either on the genetic and conservation consequences of habitat fragmentation (Mader 1984, Reh 1989) or on direct mortality caused by vehicular traffic (Case 1978, Bernadino and Dalrymple 1992, Rosen and Lowe 1994, Ashley and Robinson 1996, Gokula 1997, Clevenger et al. 2003). However, the proximate mechanisms that cause roads to influence reptile populations remain virtually unstudied. The central issue involves behavioral responses by reptiles to roads. For example, if a reptile species actively avoids roads, then highway mortality will be minimal, but local populations may be effectively fragmented by any road construction. At the other extreme, reptiles may actively seek out roads, perhaps attracted to the sun-warmed road surface for thermal reasons (Klauber 1956, Gibbons and Semlitsch 1987, Rosen and Lowe 1994, Ashley and Robinson 1996, Gokula 1997). Such a species might experience high rates of roadkill.
Information on behavioral responses of free-ranging reptiles to roads is needed to clarify these questions, but available data are limited and typically indirect. For example, radio-tracked lizards crossed roads less often than expected under null models based on road densities (Klingenböck et al. 2000, Koenig et al. 2001); and snakes are often collected by road-driving at night, with many authors speculating that the animals actively seek out roads for thermal reasons (Klauber 1956, Gibbons and Semlitsch 1987, Rosen and Lowe 1994, Ashley and Robinson 1996, Gokula 1997). Reproduction may also be affected, with nesting concentrated along forest clearings that allow sun penetration and thus warmer nests (Zuiderwijk et al. 1993, Shine et al. 2002). Such behavioral responses may be important proximate determinants of the degree to which road construction modifies population connectedness and viability.
We are not aware of any attempt to quantify direct effects of roads on reptile behavior. Therefore, we have conducted a study on the ways in which a gravel road affects red-sided garter snakes (Thamnophis sirtalis parietalis) near a communal den in central Canada. Our study aimed to answer the following questions: (1) Does the road surface differ from surrounding habitats in attributes that might plausibly affect a reptile's thermoregulatory opportunities, predation exposure, etc.? (2) Does the road affect snake behavior? Specifically, do the animals spend more or less time on the road than would be expected by chance? and (3) Does the road impede mate location?
Red-sided garter snakes are medium sized (the average snout to vent length (SVL) of adult males is 45 cm, and of females 60 cm), active, diurnal, terrestrial non-venomous snakes. In the Interlake region of central Manitoba, severely cold winters force these snakes to gather in large numbers at communal overwintering dens (Aleksiuk and Gregory 1974, Gregory and Stewart 1975, Gregory 1977). The snakes court and mate in early spring before dispersing to their summer feeding ranges (Gregory 1974). Females disperse to their summer ranges less than 3 days after emergence, but many males wait near the den for more than 2 weeks to intercept emerging females (Gregory 1974, Shine et al. 2001). Thus, sex ratios at the den in spring are heavily male biased (Gregory 1974). Males locate females by chemoreception, and use their sophisticated vomeronasal system to follow pheromonal trails laid down on the substrate by dispersing females (LeMaster et al. 2001).
In May 2003, we worked at a communal den containing more than 10 000 snakes, located 1.5 km north of the town of Inwood, and 250 m east of Highway 17 in central southern Manitoba (50°31.58'N 97°29.71'W). The area is a mosaic of aspen woodland and open pastures. At our study site, the earth has been excavated over about 3 ha to a depth of 4 m for a limestone quarry. To provide vehicular access from the grassland to the floor of the quarry, a road was constructed by piling crushed limestone boulders to provide a firm base; the surface of the road consists of compacted, pulverized limestone pebbles. The snakes emerge from small holes between the boulders beneath the road, and from adjacent areas where this material has been deposited (Fig. 1). The road is 4 m wide and runs east–west, parallel to and <5 m from the main den area from whence the snakes emerge. Thus, many snakes encounter the road during their daily activities (courtship, mating, dispersal). No vehicles use the road during the time that the snakes are active in spring (April–June), but trucks regularly use the road during the summer for quarry excavation work.
|
Fig. 1. Gravel road adjacent to a communal den of garter snakes (Thamnophis sirtalis parietalis) near Inwood, Manitoba. Note that snakes crossing the road generally do so by moving perpendicular to the main axis of the road.

|
Road Attributes
To quantify abiotic features of the habitat provided by the road, we set out six parallel transects running from south to north, perpendicular to the east–west axis of the road. Adjacent transects were separated by 6 m, and each had seven points at 1.5-m intervals. The innermost point on each transect was on the center of the road surface, the two outermost points on each end of each transect were in the surrounding grass and aspen habitat, and the remaining two points on each side, midway along the transects, were on the road but close to its edge. All points were marked with painted rocks, enabling us to remeasure attributes at the same specific locations. At each of the seven points on each of the six transects (total of 42 sampling sites), we measured vegetation cover, canopy openness, substrate reflectance, and substrate temperature.
Vegetation cover (percentage of the ground surface covered by vegetation within each half of a 1-m diameter circle around each specified point) was estimated visually.
Canopy openness was quantified by placing a 35-mm camera with a 180° hemispherical lens (f1 7.5 mm fisheye lens, Canon, Japan) on each transect point and taking a photograph pointing directly upward. The resulting photographs were scanned and specialized GLA image-analysis software was used to extract information on canopy cover and total incident daily solar radiation (Frazer et al. 1999).
Substrate reflectance was measured by holding a Nikon FM10 35-mm SLR camera 1 m above the ground pointing directly downward and recording the f-stop for correct exposure at 1/250 sec. These readings were taken only once at each site, under similar weather conditions; the data were later converted to lux by calibrating the camera against a Minolta 5 flash meter.
Substrate temperature was measured by holding a Raytek Raynger ST2L infrared thermometer 1 m above the ground, pointing directly downward. The resultant reading quantified average temperature over a 12.5-cm diameter circular area, centered on the specified transect point. Readings were taken once during each 1-h period from 0800 to 2200 h, between 21 and 24 May 2003.
To compare these attributes of a gravel (unsealed) road with those of an asphalt (bitumen) highway, we also took a smaller sample of readings on Highway 17, 1 km west of the Inwood den. The highway is 6 m wide, runs north–south, is elevated 1 m above the general ground level, and is bordered on both sides by open grassy verges 8 m wide. Vehicular traffic prevented us from measuring canopy openness at this site, and temperature readings were taken only at 0700, 1100, 1330, and 1950 h.
Behavioral Responses of Snakes to the RoadIf snakes are affected by the presence of a road, we may expect that their patterns of distribution and movement will be modified as a result. Specifically, if snakes tend to minimize the time spent on the road surface, then we predict that (a) snake densities should be lower on the road than in adjacent areas; (b) snakes approaching the road should frequently turn at an abrupt angle and either travel along the verge (parallel to the road) or move back into the surrounding habitat; and (c) snakes crossing the road should do so at an angle (perpendicular to the main axis of the road) that minimizes distance traveled on the road surface.
To test whether snake densities were lower on the road than in adjacent areas, we scored the numbers of snakes in 1-m diameter circular areas around each of our survey sites whenever we measured the abiotic variables described above. To determine whether snakes frequently turn at an abrupt angle when encountering a road, we measured the angles of movement of snakes by standing near the edge of the road and scoring snakes that were first seen >3 m from the road, and traveling towards it. We measured the angles of crossing using the same method above, but looking only at snakes that actually crossed the road. For both approaching and crossing snakes, the angle was calculated perpendicular to the main axis of the road (i.e., a snake proceeding directly across the road, at right angles to the main axis of the road, would score 0°). Because sex ratios at the den in spring are highly male biased (Gregory 1974), and female movements are influenced by the distribution of males (females actively avoid males, Shine et al. 2003), all data in this paper are based only on adult male snakes.
Ability of Trail-Following Male Snakes to Locate FemalesAdult male garter snakes at the Inwood den spend most of their time either courting females or following substrate-deposited pheromonal (lipid) trails in order to locate females (LeMaster et al. 2001). Plausibly, the crushed limestone surface of the road might influence a male's ability, or willingness, to accurately follow a female's trail. Thus, we set out experimental trails by allowing females to crawl either through grassland near the road, or across the road, and then released males at the start of these trails to examine their tactics and success rates in mate location. We ran six trials where the female trail was laid across the gravel road, and six trials where the trail was laid through grassland adjacent to the road.
Adult female snakes found being courted in the den (and thus, known to be attractive) were collected, and their unmated status confirmed by the absence of copulatory plugs (Shine et al. 2000). Each female was then placed on the ground 50 m away from the den and encouraged to crawl for 5 m either across the gravel road or through open grassland 2 m from the road. Although snakes generally avoid roads (see below), a female could easily be encouraged to travel across the road by facing her in the correct direction at the time of release. We then collected and removed the female, and placed another reproductive female (hereafter, called the “destination female”) at the end of the pheromonal trail left by the first female. To ensure that she stayed in place, the destination female was anesthetized by intramuscular injection of sodium brevital at 0.003 mL/g body mass (enough to prevent movement for >10 min). A plastic garbage can (diameter 45 cm; height 60 cm) with its base cut out was placed upside down on the ground at the place of origin of the first female's trail, and another anesthetized female (hereafter, called the “origin female”) was placed inside it, with her head protruding through a small opening cut into the rim of the garbage can at ground level, directly above the beginning of the pheromonal trail. For each trial, ten male snakes found courting in the den were then placed into the garbage can. As soon as they began actively courting the anesthetized origin female (usually within 30 sec), she was removed by pulling her headfirst through the ground-level opening. In every case, males' heads appeared at this opening <10 sec later, and they exhibited stereotypical trail-following behaviors (frequent tongue flicking to the substrate, with sweeping lateral movements of the head; LeMaster et al. 2001) as they moved along the pheromonal trail. The observer remained immobile near the garbage can, recording male behavior and scoring the elapsed time until males found the destination female (in every case, they began courting her <5 sec after first touching her body).
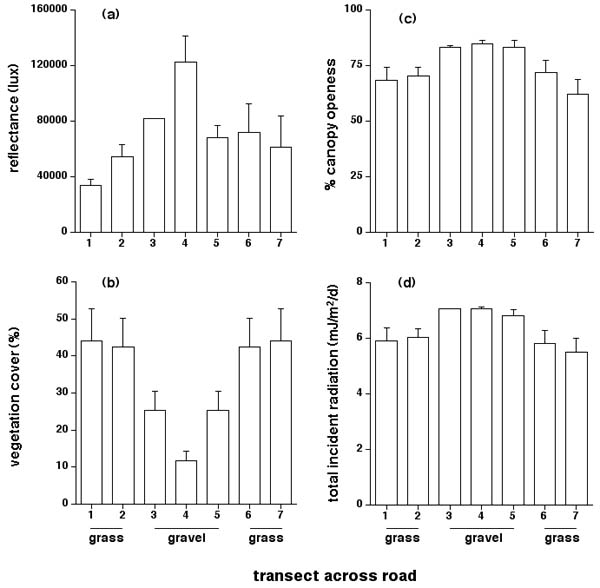
Unsurprisingly, many characteristics of the road surface differed from those along the verge or in the surrounding grassland. Figures 1 and 2b show that vegetation cover at ground level differed among sites along our transects (F6,77 = 3.60, p < 0.004; Fishers PLSD posthoc tests reveal that ground cover at site 4 in the center of the road averaged significantly lower than sites at the edges [1, 2, 6, 7]). Trees and bushes were absent from the open gravel habitat along the road center, and grass was scarce (Fig. 1). Thus, canopy openness was higher in the center of the road (Fig. 2c; F6,35 = 3.92, p < 0.005; posthoc tests show that the central sites [3, 4 and 5] differed significantly from the edge sites [1, 2, 6, 7] in most comparisons). In turn, the more open canopy resulted in higher penetration of incident solar radiation to sites near the center of the road (Fig. 2d; F6,35 = 3.46, p < 0.01; posthoc tests show that the central sites [3, 4 and 5] were significantly higher than the edge sites [1, 2, 6, 7] in most comparisons).
|
Fig. 2. Attributes of the physical environment along transects across a gravel road adjacent to a communal den of garter snakes (Thamnophis sirtalis parietalis) near Inwood, Manitoba. The graphs show mean values and associated standard errors for (a) reflectances, (b) vegetation cover on the ground, (c) canopy openness, and (d) incident solar radiation striking the ground. See text for explanation of methods. “Grass” and “gravel” refer to substrate types on different parts of the transect.
|
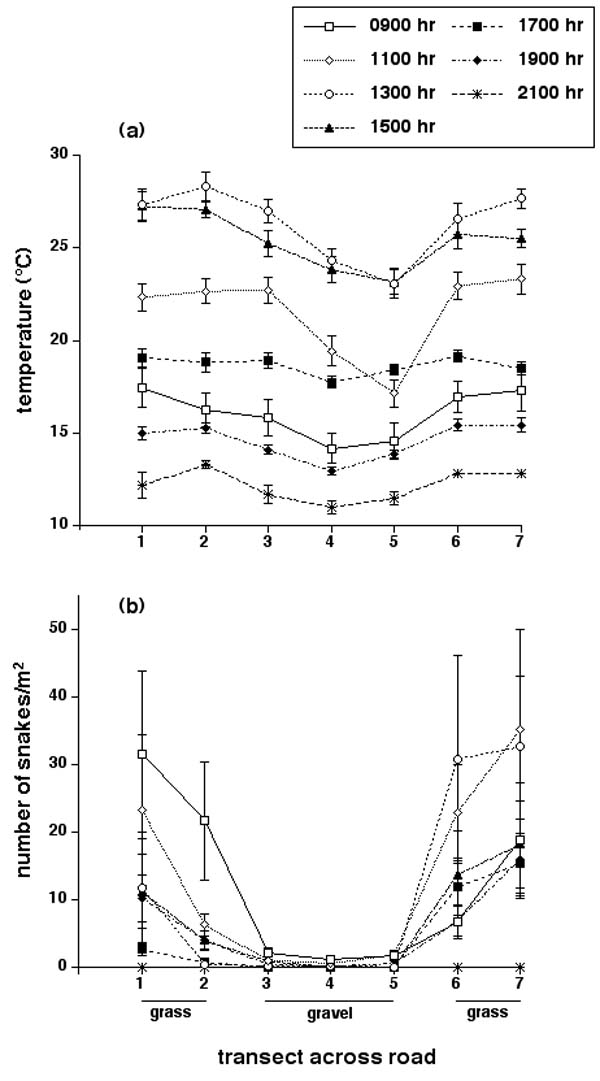
The white gravel forming the road surface was more reflective than the surrounding habitat (Figs. 1 and 2a). Substrate temperatures would be decreased by this phenomenon, but increased by the higher radiation input (above; see Fig. 2d). In practice, the former effect was stronger than the latter in terms of thermal equilibrium: thus, the road surface was cooler than its surrounds. This pattern was evident throughout the day, although most pronounced in the mid-morning as the ground surface warmed (Fig. 3a). Thermal differentials were minor in the evening (Fig. 3a). Repeated-measures ANOVA on the total data set (with sampling time as the repeated measure, site within transect as the factor, and substrate temperature as the dependent variable) showed that temperatures not only varied through time during the day (F15,510 = 318.98, p < 0.0001) and differed among sites (F6,34 = 11.83, p < 0.0001) but that the spatial heterogeneity of temperatures changed through time (interaction, F90,510 = 2.60, p < 0.0001). The significant interaction term reflects the fact that, although average temperatures were always lower on the road than in the adjacent grassland, this disparity was greater during the morning than in the late afternoon or evening (Fig. 3a).
|
Fig. 3. Effects of time of day on (a) substrate temperatures and (b) numbers of snakes along transects across a gravel road near a communal snake den. Graphs show mean values and associated standard errors.
|
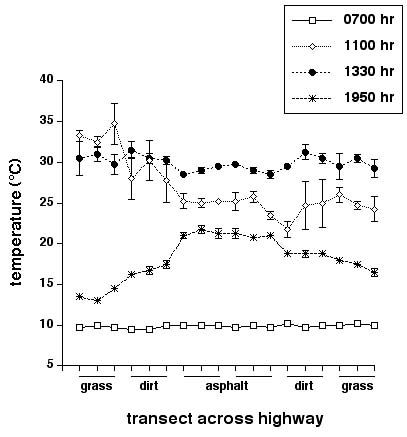
A smaller data set on habitat attributes across a nearby asphalt highway revealed a different pattern. No snakes were seen within our sample plots either on the highway or beside it, during our four sampling periods. Vegetation cover was bimodal: 100% on the grassy verge and 0% on the dirt and asphalt. As for the gravel road, reflectances were higher on the road surface (mean = 81 920 lux) than on the surrounding grassy verge (dirt = 69 973; grass = 27 306 lux; ANOVA with substrate type as the factor, F2,57 = 87.69, p < 0.0001, all posthoc comparisons have p < 0.05). However, the magnitude of these disparities was lower than for the gravel road, reflecting the much higher reflectance of the white gravel than the dark asphalt surface (120 000 vs. 80 000 lux). Substrate temperatures at the highway site showed a more dramatic and complex shift during the day than was the case for the gravel road (Fig. 4). Temperatures were uniformly low (around 10°C) early in the morning and uniformly high (around 30°C) in the early afternoon. Heating in the morning was highly asymmetric, reflecting greater sun exposure on the east-facing verge than on the western side. Throughout these daytime samples, average substrate temperatures were similar on the road surface to those on the surrounds. After sunset, however, the asphalt surface of the road was substantially (4–9°C) warmer than other parts of the transect (1950 h sample in Fig. 4: ANOVA for data taken at this time have location effect, F17,54 = 50.78, p < 0.0001, posthoc tests have all asphalt sites significantly warmer than all other sites).
|
Fig. 4. Temperatures on transects across an asphalt highway near Inwood, Manitoba at four times of day. Graph shows mean values and associated standard errors for each time period based on four replicates at each of 18 equally spaced points along transects perpendicular to the highway.
|
Behavioral Responses of Snakes to the Road
Quadrat counts consistently showed much lower numbers of snakes present on the road surface than beside the road, throughout the day (Fig. 3b). Repeated-measures ANOVA on the total data set (with sampling time as the repeated measure, site within transect as the factor, and snake densities as the dependent variable) showed that numbers of snakes varied through time during the day (F15,420 = 3.03, p < 0.0001) and differed among sites (F6,28 = 4.08, p < 0.005) but that the spatial location of snakes remained consistent through time (interaction, F90,420 = 1.14, p = 0.20). The non-significant interaction term reflects the fact that numbers of snakes were always lower on the road than in the adjacent grassland (Fig. 3b).
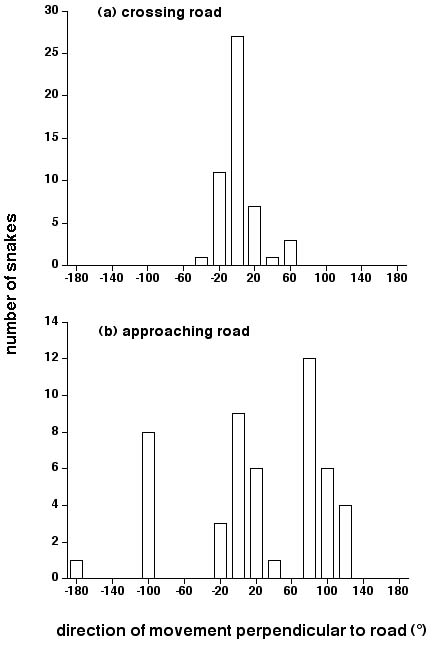
Direct observations of moving snakes showed that animals crossing the road usually did so by the shortest route, perpendicular to the main axis of the road (0° in Fig. 5a). Snakes observed as they approached the road either crossed directly (as above, 0°) or turned and moved along the verge parallel to the main axis of the road (thus, generally at +90 or -90°; see Fig. 5b). In both cases, contingency-table analysis comparing the numbers of snakes in each 20° segment to a random model of equal probabilities, strongly rejected the null hypothesis (for road crossing, χ2 = 277.6, 17 df, p < 0.0001; for approach, χ2 = 110.20, 17 df, p < 0.001). Using the same data, we also conducted tests of more specific hypotheses. The number of road-crossing males traveling within the 20° segment centered on 0° (directly perpendicular to the main axis of the road) was significantly higher than the numbers in adjacent segments either to the left or right (χ2 =7.41, 1 df, p < 0.001; χ2 = 4.83, 1 df, p < 0.05). Similarly, directions of travel for snakes crossing the road differed significantly from those approaching the road (comparing Figs. 5a vs. 5b; excluding segments where no snakes were recorded, χ2 = 48.65, 10 df, p < 0.0001).
|
Fig. 5. Direction of movement of garter snakes (Thamnophis sirtalis parietalis) with respect to the main axis of the gravel road. The lower graph shows data for snakes scored as they approached within 1 m of the road, coming from the adjacent den. The upper graph shows data for snakes scored as they actually crossed the road. For both approaching and crossing snakes, the angle was calculated perpendicular to the main axis of the road (i.e., a snake proceeding directly across the road, at right angles to the main axis of the road, would score 0°).
|
Ability of Trail-Following Male Snakes to Locate Females
The numbers of males finding the female within 5 min (out of ten males released in the garbage can for each trial) averaged 4.83 (SD = 3.37) for grassland trials and 1.17 (SD = 1.47) for road trials. One-factor ANOVA showed that this difference was statistically significant (using trials as replicates, F1,10 = 5.96, p < 0.04). The mean times for successful males to find the destination female did not differ between the two sets of trials, with successful males in grassland trials finding the female in 141.92 (SD = 81.39) sec, whereas successful males in the road trials took slightly but not significantly longer (using individual males as replicates to maximize statistical power, 151.43 ± 66.31 sec; F1,31 = 0.08, p = 0.78). Thus, fewer males relocated the female if her trail lay across the road; but those that succeeded, did so after similar periods of searching.
The mate-searching males following trails in grassland exhibited classic trail-following behavior, with heads close to the ground and frequent tongue flicking. When the trail intercepted the road, all males paused at the verge rather than crossing. Some males then abandoned the trail and dispersed in other directions away from the road; others ceased tongue flicking and instead elevated their head and forebodies in an apparent attempt to obtain visual cues to the female's location. Males that crossed the road did so quickly; only two of the males showed tongue flicking to the road surface, in contrast to all males following trails in grassland. All males resumed tongue flicking the substrate as soon as they reached vegetation on the other side of the road. One of the two males that tongue flicked the road surface recoiled and gaped, perhaps due to adhesion of dust particles to his tongue tips.
Although the gravel road near the Inwood den is small and had no vehicular traffic during our study period, it nonetheless (a) comprised a very distinctive habitat in terms of several abiotic variables and (b) significantly modified the distributions, movement patterns, and mate-location abilities of snakes. Some of the effects that we documented are intuitively obvious but have rarely been quantified; others proved to be surprising (at least to us). Below, we consider first the physical changes wrought by roads, and then the behavioral modifications that these changes induced in the snakes.
Our data on the physical attributes of sites across roads point to complex effects of combinations of variables. Most obviously, a road comprises a linear expanse of habitat with atypically low vegetation cover and thus, a relatively open canopy that allows high penetration of solar radiation (Fig. 2). Intuition would thus suggest that the road surface would be warmer than nearby, more shaded areas. In practice, the reverse was true, at least on the gravel road; the disparity sometimes exceeded 5°C (Fig. 3a). The explanation for this paradox is that the white gravel on the road surface is more reflective than the surrounding grassland (Fig. 2a). The blacktop asphalt highway presented a very different thermal situation: temperatures on the road surface were similar to those on the verges for most of the diel cycle, but the greater thermal inertia of the asphalt resulted in a substantial thermal differential in the evening, as the surrounding habitat cooled more rapidly than the road (Fig. 4). Thus, the thermal differential between the road and its surrounds sometimes exceeded 5°C on both roads, but the patterns differed dramatically both in terms of the direction of the difference (asphalt road hotter than verges, gravel road cooler) and the time of greatest thermal differential (morning for the gravel road, evening for the asphalt road).
Spatial heterogeneity in habitat temperatures strongly influences movements and habitat selection by snakes (Huey et al. 1989, Webb and Shine 1998, Blouin-Deemers and Weatherhead 2001). Given the magnitude of the thermal differentials that we measured between adjacent sites (Figs. 3 and 4), it seems likely that snakes could respond to such variation. Many authors have suggested that snakes are attracted to sun-warmed road surfaces, and that this thermally driven process contributes to their high vulnerability to vehicular traffic (e.g., Gibbons and Semlitsch 1987, Ashley and Robinson 1996). Our data paint a more complex picture. High evening temperatures may well attract snakes to asphalt roads, especially in areas with high diel variation in ambient temperatures. However, at least some gravel roads are cooler, not warmer, than natural habitats, and thus do not induce road-basking behavior. Information on the thermal profiles of dark vs. light-colored gravel roads would be of great interest in this respect; it seems clear that reflectivity of the road surface may be a more important determinant of thermal regimes than has generally been appreciated.
Snakes at the Inwood den clearly avoided the road surface, but the causal mechanisms behind these avoidance reactions are unclear: they may represent direct responses to abiotic stimuli such as temperature, or more complex responses that have evolved to enhance organismal fitness (e.g., avoid open areas because of higher risk from predation). Regardless, the magnitude of the effect is striking: numbers of snakes on the road surface averaged 2% of those on the verge, only 3 m away (Fig. 3b). Similarly, the angles at which snakes crossed the road were remarkably consistent, and almost always very close to the shortest possible distance (Fig. 5a). Lastly, snakes approaching the road typically either went straight across it, or turned to move parallel to the road's edge, along the verge (Fig. 5b). All these patterns indicate strong avoidance of the road surface. Such behavior may result in considerable decreases in rates of mortality caused by vehicular traffic (Dodd 1987, Krivda 1993, Shine and Mason 2001).
Pheromonal trail-following using the vomeronasal system plays a central role in the mating systems of most or all snake species (e.g., Gillingham 1987, Mason et al. 1989, Mason 1992, Chiaraviglio and Briguera 2001). Indeed, a review of mating systems in snakes concluded that “pheromonal trails may be the only class of ‘distance’ cues that snakes regularly use to detect the moderate to distant physical proximity of conspecifics” (Duvall et al. 1992, p. 324). Thus, any anthropogenic modification that impairs pheromonal trail-following ability may significantly affect mating systems of these animals. Our manipulative experiments provide strong evidence that male garter snakes were unable or unwilling to follow pheromonal trails of reproductive females across the gravel road. Given the high population densities of garter snakes near the Manitoba dens, such effects do not threaten population viability: even if trail following is disrupted by roads, most females will encounter numerous males. However, many snake species—including many threatened taxa—occur at very low population densities. Although such low densities are a natural feature of population biology for many predators at high trophic levels such as snakes, taxa that are already at risk may be vulnerable to any disturbance that reduces rates of mate location.
Impaired mate-location abilities caused by the presence of roads may have diverse consequences. Most obviously, some potentially reproductive females may be unable to breed because they are not inseminated. At a more subtle level, the viability of a female's offspring may be reduced if she is unable to mate with more than one male (Madsen et al. 1992). Such genetic benefits of multiple mating are likely to be most extreme in small inbred populations, exactly the ones for which roads may pose barriers to mate location. Indeed, such barriers may tend to reduce effective population sizes by dividing previously continuous populations into smaller fragments, further challenging their long-term viability. Lastly, impediments to the effectiveness of trail following may result in males traveling further and longer to locate a female, and thus being exposed to numerous sources of mortality (Aldridge and Brown 1995, Bonnet et al. 1999).
The substantial thermal differences between our gravel road and the adjacent asphalt highway (Figs. 3 vs. 4) suggest that snakes may respond differently to these two types of roads. Highways differ in other important respects also, notably in their width and in the frequency of vehicular traffic. Diel cycles in the thermal profiles of the highway suggest that nocturnally active snakes may be at particular risk, because the road is warmer than the surrounding grassland early in the evening (Fig. 4) and thus may be thermally attractive to snakes at this time. Given such differences, further research on the responses of snakes to asphalt vs. gravel roads is needed.
Our data suggest that the proliferation of roads—including small gravel tracks and larger highways—may directly affect snake behaviors and mating systems. This is a worrying conclusion, because of the enormous proliferation of roads in the landscape over recent decades. For example, Riitters and Wickham's (2003) analysis of North American ecosystems estimated that 20% of the total land area was <127 m from a road. In combination with other data demonstrating complex and sometimes dramatic effects of roads on other important abiotic and biotic factors that may affect population viability, our study suggests that further research is urgently needed on the ways in which roads influence snake populations.
Responses to this article are invited. If accepted for publication, your response will be hyperlinked to the article. To submit a comment, follow this link. To read comments already accepted, follow this link.
We thank Al and Gerry Johnson for help and encouragement, and the Manitoba Department of Natural Resources (especially Dave Roberts) for permits. Financial support was provided by the Australian Research Council (to RS), by a National Science Foundation National Young Investigator Award (IBN-9357245), and by the Whitehall Foundation (W95-04) to RTM. Research was conducted under the authority of Oregon State University Institutional Animal Care and Use Committee Protocol No. LAR-1848B. All research was conducted in accordance with the US Public Health Service “Policy on Humane Care and Use of Laboratory Animals” and the National Institutes of Health “Guide to the Care and Use of Laboratory Animals.”
Aldridge, R. D., and W. S. Brown. 1995. Male reproductive cycle, age at maturity, and cost of reproduction in the timber rattlesnake (Crotalus horridus). Journal of Herpetology 29:399–407.
Aleksiuk, M., and P. T. Gregory. 1974. Regulation of seasonal mating behavior in Thamnophis sirtalis parietalis. Copeia 1974:682–689.
Alexander, S. M., and N. M. Waters. 2000. The effects of highway transportation corridors on wildlife: a case study of Banff National Park. Transportation Research Part C - Emerging Technologies 8:307–320.
Ashley, E. P., and J. T. Robinson. 1996. Road mortality of amphibians, reptiles and other wildlife on the Long Point causeway, Lake Erie, Ontario. Canadian Field Naturalist 110:403–412.
Bernadino, F. S., and G. H. Dalrymple. 1992. Seasonal activity and road mortality of the snakes of the Pa-hay-okee wetlands of Everglades National Park, USA. Biological Conservation 62:71–75.
Blouin-Demers, G., and P. J. Weatherhead. 2001. An experimental test of the link between foraging, habitat selection and thermoregulation in black rat snakes Elaphe obsoleta obsoleta. Journal of Animal Ecology 70:1006–1013.
Bonnet, X., G. Naulleau, and R. Shine. 1999. The dangers of leaving home: dispersal and mortality in snakes. Biological Conservation 89:39–50.
Case, R. M. 1978. Interstate highway road-killed animals: a data source for biologists. Wildlife Society Bulletin 6:8–13.
Chiaraviglio, M., and V. Briguera. 2001. Participation of chemical cues in conspecific detection and discrimination in Boa constrictor (Serpentes: Boidae). Gayana Zoologica 65:5–10.
Clevenger, A. P., B. Chruszcz, and K. E. Gunson. 2001. Highway mitigation fencing reduces wildlife–vehicle collisions. Wildlife Society Bulletin 29:646–653.
Clevenger, A. P., B. Chruszcz, and K. E. Gunson. 2003. Spatial patterns and factors influencing small vertebrate fauna road-kill aggregations. Biological Conservation 109:15–26.
Dodd, C. K. 1987. Status, conservation, and management. Pages 478–513 in R. A. Seigel, J. T. Collins, and S. S. Novak, editors. Snakes: ecology and evolutionary biology. MacMillan, New York, New York, USA.
Duvall, D., G. Schuett, and S. J. Arnold. 1992. Pitviper mating systems: ecological potential, sexual selection and microevolution. Pages 321–336 in J. A. Campbell and E. D. J. Brodie, editors. Biology of the pitvipers. Selva, Tyler, Texas, USA.
Fehlberg, U. 1994. Ecological barrier effects of motorways on mammalian wildlife—an animal protection problem. Deutsche Tierarztliche Wochenschrift 101:125–129.
Forman, R. T. T. 2000. Estimate of the area affected ecologically by the road system in the United States. Conservation Biology 14:31–35.
Frazer, G. W., C. D. Canham, and K. P. Lertzman. 1999. Gap Light Analyzer (GLA), Version 2.0: imaging software to extract canopy structure and gap light transmission indices from true-color fisheye photographs, users manual and program documentation. Simon Fraser University, Burnaby, British Columbia, Canada.
Gibbons, J. W., and R. D. Semlitsch. 1987. Activity patterns. Pages 396–421 in R. A. Seigel, J. T. Collins, and S. S. Novak, editors. Snakes: ecology and evolutionary biology. MacMillan, New York, New York, USA.
Gokula, V. 1997. Impact of vehicular traffic on snakes in Mudumalai Wildlife Sanctuary. Cobra 27:26–30.
Gillingham, J. C. 1987. Social behavior. Pages 184–209 in R. A. Seigel, J. T. Collins, and S. S. Novak, editors. Snakes: ecology and evolutionary biology. MacMillan, New York, New York, USA.
Gregory, P. T. 1974. Patterns of spring emergence of the red-sided garter snake (Thamnophis sirtalis parietalis) in the Interlake region of Manitoba. Canadian Journal of Zoology 52:1063–1069.
Gregory, P. T. 1977. Life-history parameters of the red-sided garter snake (Thamnophis sirtalis parietalis) in an extreme environment, the Interlake region of Manitoba. National Museum of Canada, Publications in Zoology 13:1–44.
Gregory, P. T., and K. W. Stewart. 1975. Long-distance dispersal and feeding strategy of the red-sided garter snake (Thamnophis sirtalis parietalis) in the Interlake of Manitoba. Canadian Journal of Zoology 53:238–245.
Groot Bruinderink, G. W. T. A., and E. Hazebroek. 1996. Ungulate traffic collisions in Europe. Conservation Biology 10:1059–1067.
Huey, R. B., C. R. Peterson, S. J. Arnold, and W. P. Porter. 1989. Hot rocks and not-so-hot rocks: retreat-site selection by garter snakes and its thermal consequences. Ecology 70:931–944.
Jones, M. E. 2000. Road upgrade, road mortality and remedial measures: impacts on a population of eastern quolls and Tasmanian devils. Wildlife Research 27:289–296.
Klauber, L. M. 1956. Rattlesnakes. Their habits, life histories, and influence on mankind. University of California Press, Berkeley, California, USA.
Klingenböck, A., K. Osterwalder, and R. Shine. 2000. Habitat use and thermal biology of the “land mullet” Egernia major, a large scincid lizard from remnant rainforest in southeastern Australia. Copeia 2000:931–939.
Koenig, J., R. Shine, and G. Shea. 2001. The ecology of an Australian reptile icon: how do bluetongue lizards (Tiliqua scincoides) survive in suburbia? Wildlife Research 28:215–227.
Krivda, W. 1993. Road kills of migrating garter snakes at The Pas, Manitoba. Blue Jay 51:197–198.
LeMaster, M. P., I. T. Moore, and R. T. Mason. 2001. Conspecific trailing behavior of red-sided garter snakes, Thamnophis sirtalis parietalis, in the natural environment. Animal Behaviour 61:827–833.
Lodé, T. 2000. Effect of a motorway on mortality and isolation of wildlife populations. Ambio 29:163–166.
Mader, H-J. 1984. Animal habitat isolation by roads and agricultural fields. Biological Conservation 29:81–96.
Madsen, T., R. Shine, J. Loman, and T. Håkansson. 1992. Why do female adders copulate so frequently? Nature 335:440–441.
Mason, R. T. 1992. Reptilian pheromones. Pages 114–228 in C. Gans and D. Crews, editors. Biology of the Reptilia, vol. 18. Hormones, brains and behavior. University of Chicago Press, Chicago, Illinois, USA.
Mason, R. T., H. M. Fales, T. H. Jones, L. K. Pannell, J. W. Chinn, and D. Crews. 1989. Sex pheromones in snakes. Science 245:290–293.
May, S. A., and T. W. Norton. 1996. Influence of fragmentation and disturbance on the potential impact of feral predators on native fauna in Australian forest ecosystems. Wildlife Research 23:387–400.
Nielsen, C. K., R. G. Anderson, and M. D. Grund. 2003. Landscape influences on deer–vehicle accident areas in an urban environment. Journal of Wildlife Management 67:46–51.
Putman, R. J. 1997. Deer and road traffic accidents: options for management. Journal of Environmental Management 51:43–57.
Rea, R. V. 2003. Modifying roadside vegetation management practices to reduce vehicular collisions with moose Alces alces. Wildlife Biology 9:81–91.
Reh, W. 1989. Investigations into the influence of roads on the genetic structure of populations of the common frog Rana temporaria. Pages 101–103 in T. E. S. Langton, editor. Amphibians and roads: Proceedings of the Toad Tunnel Conference. Rendsburg, Federal Republic of Germany, 7-8 January 1989. ACO Polymer Products Ltd., Bedfordshire, UK.
Ries, L., D. M. Debinski, and M. L. Wieland. 2001. Conservation value of roadside prairie restoration to butterfly communities. Conservation Biology 15:401–411.
Riitters, K. H., and J. D. Wickham. 2003. How far to the nearest road? Frontiers in Ecology and the Environment 1(3):125–129.
Rosen, P. C., and C. H. Lowe. 1994. Highway mortality of snakes in the Sonoran desert of southern Arizona. Biological Conservation 68:143–148.
Seabrook, W. A., and E. B. Dettmann. 1996. Roads as activity corridors for cane toads in Australia. Journal of Wildlife Management 60:363–368.
Shine, R., E. G. Barrott, and M. J. Elphick. 2002. Some like it hot: effects of forest clearing on nest temperatures of montane reptiles. Ecology 83:2808–2815.
Shine, R., M. J. Elphick, P. S. Harlow, I. T. Moore, M. P. LeMaster, and R. T. Mason. 2001. Movements, mating and dispersal of red-sided gartersnakes from a communal den in Manitoba. Copeia 2001:82–91.
Shine, R., and R. T. Mason. 2001. Serpentine cross-dressers. Natural History 110:56–61.
Shine, R., M. M. Olsson, and R. T. Mason. 2000. Chastity belts in gartersnakes: the functional significance of mating plugs. Biological Journal of the Linnean Society 70:377–390.
Shine, R., B. Phillips, T. Langkilde, D. Lutterschmidt, H. Waye, and R. T. Mason. 2004. Mechanisms and consequences of sexual conflict in garter snakes (Thamnophis sirtalis, Colubridae). Behavioral Ecology: in press.
Stiles, J. H., and R. H. Jones. 1998. Distribution of the red imported fire ant, Solenopsis invicta, in road and powerline habitats. Landscape Ecology 13:335–346.
Waring, G. H., J. L. Griffis, and M. E. Vaughn. 1991. White-tailed deer roadside behavior, wildlife warning reflectors, and highway mortality. Applied Animal Behavior Science 29:215–223.
Webb, J. K., and R. Shine. 1998. Thermoregulation by a nocturnal elapid snake (Hoplocephalus bungaroides) in south-eastern Australia. Physiological Zoology 71:680–692.
Zuiderwijk, A., G. Smit, and H. Van Den Bogert. 1993. Man-made hatcheries for grass snakes (Natrix natrix L. 1758): a simple method for grass snake conservation. Mertensiella 3:227–234.
Address of Correspondent:
Richard Shine
Biological Sciences A08,
University of Sydney,
NSW 2006
Australia
Phone: 612-9351-3772
Fax: 612-9351-5609
rics@bio.usyd.edu.au
![]()