|
|
|
Copyright © 2001 by the author(s). Published here under license by The Resilience Alliance.
The following is the established format for referencing this article:
Bélisle, M. and C. C. St. Clair. 2001. Cumulative effects of barriers on the movements of forest birds. Conservation Ecology 5(2): 9. [online] URL: http://www.consecol.org/vol5/iss2/art9/
A version of this article in which text, figures, tables, and appendices are separate files may be found by following this link.
Report Cumulative Effects of Barriers on the Movements of Forest Birds Marc Bélisle and Colleen Cassady St. Clair
University of Alberta
- Abstract
- Introduction
- Methods
- Results
- Discussion
- Responses To this Article
- Acknowledgments
- Literature Cited
Although there is a consensus of opinion that habitat fragmentation has deleterious effects on animal populations, primarily by inhibiting dispersal among remaining patches, there have been few explicit demonstrations of the ways by which degraded habitats actually constrain individual movement. Two impediments are primarily responsible for this paucity: it is difficult to separate the effects of habitat fragmentation (configuration) from habitat loss (composition), and conventional measures of fragmented habitats are assumed to be, but probably are not, isotropic. We addressed these limitations by standardizing differences in forest cover in a clearly anisotropic configuration of habitat fragmentation by conducting a homing experiment with three species of forest birds in the Bow Valley of Banff National Park, Canada. Birds were translocated (1.2–3.5 km) either parallel or perpendicular to four/five parallel barriers that are assumed to impede the cross-valley travel of forest-dependent animals. Taken together, individuals exhibited longer return times when they were translocated across these barriers, but differences among species suggest a more complex interpretation. A long-distance migrant (Yellow-rumped Warbler, Dendroica coronata) behaved as predicted, but a short-distance migrant (Golden-crowned Kinglet, Regulus satrapa) was indifferent to barrier configuration. A resident (Red-breasted Nuthatch, Sitta canadensis) exhibited longer return times when it was translocated parallel to the barriers. Our results suggest that an anisotropic arrangement of small, open areas in fragmented landscapes can have a cumulative barrier effect on the movement of forest animals, but that both modelers and managers will have to acknowledge potentially counterintuitive differences among species to predict the effect that these may have on individual movement and, ultimately, dispersal.
KEY WORDS: Banff National Park, Golden-crowned Kinglet, Red-breasted Nuthatch, Yellow-rumped Warbler, barriers, connectivity, corridor, forest cover, fragmentation, habitat loss, movement of forest birds, roads.
Published: December 11, 2001
Much recent attention has focused on forest bird movement in fragmented habitats because of the many processes by which fragmentation is expected to influence population viability. As examples, isolation of habitat patches is purported to disrupt dispersal (Matthysen et al. 1995, Machtans et al. 1996, Verhulst et al. 1997), reduce patch colonization (Verboom et al. 1991, Villard et al. 1995, van Langevelde 2000), and constrain home range and territory placement (Rail et al. 1997, Grubb and Doherty 1999, Hinsley 2000). Despite the theoretical plausibility of these effects, empirical study under natural field conditions remains limited. Such study is difficult partly because the effects of habitat fragmentation are often confounded by the effects of habitat loss, which are generally more severe (Fahrig 1997, Bender et al. 1998, Villard et al. 1999). Put another way, it is difficult to assess the respective influence of landscape composition and configuration on the movements of forest birds because these factors are often strongly intercorrelated (Desrochers et al. 1999, Bélisle et al. 2001). For instance, both the number of forest patches and their proximity to one another are typically inversely correlated with the amount of forest cover remaining in both agricultural and timber-harvested landscapes (Haines-Young and Chopping 1996, Schumaker 1996, Hargis et al. 1998; for a review, see Tischendorf 2001).
Another potential complication in separating the effects of landscape composition and configuration, particularly in the context of animal movement, results from assuming that landscape configuration can be described with isotropic metrics. Isotropic variables must provide the same value, regardless of the direction in which they are measured, consistent with the expectations of diffusion-like or distance-dependent dispersal models (Turchin 1998). However, such an assumption does not acknowledge either the axis or direction of movement of individuals in a fragmented landscape, and this can fundamentally affect the importance of landscape configuration on both predicted and actual rates of movement (Bélisle et al. 2001). As an illustrative example, a program like FRAGSTATS (McGarigal and Marks 1995) applied to a landscape composed of alternating patches of forest and parallel strip-cuts would provide as output a single value per configuration variable that is intended to describe its functional connectivity (sensu Tischendorf and Fahrig 2000). Yet a forest bird moving through this landscape would obviously be differentially affected if it were traveling parallel vs. perpendicular to the cuts. Such anisotropy, in which the effect of landscape configuration on movement is clearly dependent on the direction of measurement, will presumably result in differential movement rates of dispersing individuals corresponding to different axes in a particular landscape. If these differences are large, they will have considerable implications for the estimation of emigration and immigration parameters in population models. Thus, understanding the effect of landscape anisotropy on animal movement very much invites further investigation.
Anisotropic functional connectivity will characterize landscapes where large or multiple linear barriers run closely parallel to one another. Such conditions exist in parts of the Canadian Rocky Mountains comprising much protected habitat that is dissected by both natural and artificial barriers (Page et al. 1996, Wilcox et al. 1998), which generally parallel valley bottoms. In management terms, these parallel barriers may create a cumulative effect to the movement of forest-dwelling animals disproportionate to the area that they occupy and that exceeds the sum of their individual effects. Among the artificial barriers, roads have been the subject of considerable recent study here and elsewhere, and are known to profoundly impede the movement of many animal taxa (Forman and Alexander 1998, Clevenger and Waltho 2000, Trombulak and Frissell 2000). Together with other linear features like pipe, utility, and railway lines, as well as natural barriers like rivers, these valley-bottom barriers are thus likely to make cross-valley travel considerably more difficult than travel parallel to the valley bottom for forest-dwelling species. This may be true even if the actual reduction in forest cover in a trajectory that runs perpendicular, rather than parallel, to these barriers is small. We can profitably test the effect of such anisotropy on forest birds because their young disperse at a scale comparable to the size and proximity of these barriers, yet their movements in Banff and elsewhere appear to be influenced by gaps in forest cover as small as 50 m (Desrochers and Hannon 1997, St. Clair et al. 1998; C. C. St. Clair, unpublished data). We would here expect that forest birds should travel more easily, and thus quickly, along than across the valley floors.
We tested this prediction during the breeding season in Banff National Park, Alberta, Canada, using homing trials (sensu Bélisle et al. 2001) that capitalize on the relatively consistent motivation that relocated territorial birds must return home. More specifically, homing trials consisted of translocating territorial, mated males of three forest-dwelling species either across a valley floor (such that birds would repeatedly encounter movement barriers), or along it (so that birds should actually be channeled by them), to delineate in both cases a predictable direction of movement. The three species used in the experiment were the Red-breasted Nuthatch (Sitta canadensis, a facultative year-round resident), the Golden-crowned Kinglet (Regulus satrapa, a short-distance migrant), and the Yellow-rumped Warbler (Dendroica coronata, a long-distance migrant). We chose these three species because most, if not all, of their activities are restricted to forest habitats or their edges, and because they defend breeding territories of similar sizes and are thus likely to perform daily movements at comparable spatial scales (Ingold and Galati 1997, Hunt and Flaspohler 1998, Ghalambor and Martin 1999). Our choice was further motivated by the fact that their sensitivity to movement barriers caused by open areas had already been studied (Desrochers and Hannon 1997) and because they present different migratory life histories that could influence their response to cumulative barrier effects. There is a common expectation that migratory habits may influence sensitivity to habitat fragmentation (Hunter 1992, Wiens 1994), yet there are no comparisons and direct tests of this effect. As a response variable, we measured the duration with which translocated birds returned to their territories, predicting that all species would take longer when relocated across barriers.
Fieldwork was conducted within a 30 km long section of the Bow Valley immediately west of the start of the Bow Valley Parkway in Banff National Park (51° 10’ N, 115° 40’ W), Alberta, Canada. The treed portion of the valley spans 3-5 km in width and consists mainly of mature lodgepole pine (Pinus contorta) forest with smaller stands of white spruce (Picea glauca) and trembling aspen (Populus tremuloides). A portion of the forests located on the north slope of the valley (i.e., north of the secondary road, Fig. 1) burned in 1993, but not all of the trees were affected by the fire and most burned trees are still standing. Five main potential barriers to movements of forest birds run parallel to one another along the floor of the valley (from south to north in most parts of the valley; see Fig. 1): (1) the Trans Canada Highway, a four-lane highway (width ~60–100 m, including verges) that is the major east–west transportation corridor through Alberta; (2) the Bow River (width ~40–60 m); (3) the Canadian Pacific Railway mainline (width ~20–30 m); (4) a utility line (width ~20 m); and (5) the Bow Valley Parkway, a two-lane, secondary road (width ~20 m, including verges). Amid this general configuration, there were several anomalies. First, although the utility line is adjacent to the railway along most (about 70%) of its length, it runs north of the secondary road in the eastern part of our study area. Two stretches (0.6 and 1.3 km) of the highway are split into two arms by a forested median ~30 m wide, and some stretches of the river contain two to three arms, which have approximately the same width as the main course. Other open habitats include shrubby and grassy meadows, open bogs, ponds, and streams. Because these open habitats and structures were sometimes adjacent to one another, the actual width of putative barriers and the amount of nonforested habitat that they created varied with location and was thus calculated for each bird separately, as we will decribe. Translocation trials were conducted from 8 May to 29 June 2000, and spanned the respective nest building, egg laying, and egg incubation phases of the bird species under study. Subject to this constraint, we tried to balance translocations among species and between treatments on any given date. Because trials were dispersed in time, we probably avoided introducing confounding effects linked to the birds' breeding phenology or weather conditions.
|
Fig. 1. Two views of the Bow Valley of Banff National Park, Canada, and its barriers. Panel A (photo by C. C. St. Clair) depicts the parallel vs. perpendicular treatments to which translocated birds were exposed in a west-facing photo taken just west of the study site. Panel B (photo by M. Hebblewhite) shows a portion of the study area in winter when the contrast between forested and nonforested habitat is more apparent. This east-facing photo was taken about 3 km east of the western limit of the study area and shows the locations of the five barriers. Only the cleared slopes of its verges are visible for the highway.

|
Translocation procedure
We translocated territorial, mated male Red-breasted Nuthatches (Sitta canadensis, facultative year-round residents), Golden-crowned Kinglets (Regulus satrapa, short-distance migrants), and Yellow-rumped Warblers (Dendroica coronata, long-distance migrants). Females were not used because their attachment to a given territory may be quite variable, and because they bear the stress of egg laying and assume most, if not all, of the incubation. We considered a male to be mated if a female was observed in its territory on the day of the capture. Birds were caught shortly after dawn (before 0900 Mountain Standard Time) with mist nets. We attracted birds in the nets with playbacks of their territorial calls or songs. Once caught, birds were leg-banded with a unique combination of 2-4 color bands to allow recognition by sight. We translocated birds by car to limit time in captivity to < 40 min.
Seven birds of each of the three species were translocated in each of the two treatments, for a total of 42 birds. All birds were caught along a two-lane, secondary road (the Bow Valley Parkway) and were translocated either along or across the valley so that birds would be expected to be either channeled or impeded, respectively, by the series of linear features paralleling the valley bottom. Birds that were translocated along the valley were released between the south-facing mountain range and secondary road or between the latter and the utility line, depending on which side of the secondary road they were captured. We needed to capture birds on both sides of the secondary road to obtain adequate replication for each of the target species. Birds that were translocated across the valley faced either four or five barriers on their homing journey, depending on whether they had been captured between the utility line and the secondary road (14 birds) or between the latter and the south-facing mountain range (28 birds), respectively. Release sites were chosen to match relocation distance across species and treatments, but with the constraints imposed by road access. All birds were released in large forest tracts. Translocation distances were measured using the position of the capture and release sites obtained with a Trimble Geoexplorer 3™ Global Positioning System receiver with an accuracy of ± 10 m. Translocation distances ranged from 1204 to 3530 m (Table 1). These distances should be large enough to provide relevant information on landscape-scale movements, but small enough to avoid confounding effects due to the birds’ navigational ability. This range of distances is also comparable to the known natal dispersal distances of small forest birds (Anders et al. 1998, Paradis et al. 1998, Vega Rivera et al. 1998).
|
Table 1. Description of the 42 landscape sections (seven sections per treatment per species) used in translocation trials along and across potential barriers to movement in the Bow River Valley bottom, Banff National Park, Canada, in 2000.
b Forest cover was estimated in 5-m grid cells along the capture-to-release line as well as in the grid cells contained in rectangles radiating 400 m from both the line and from its start and end points. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
We allowed up to 10 days (256 h post-release) for the translocated birds to return to their respective territory. Each day, two observers checked for the presence of the birds by patrolling territories during a 30-min visit while broadcasting the bird species' territorial calls for a duration of 1-2 min every 5 min. On the translocation day, we first visited capture sites at 4.5 h post-release. On the following day, birds were checked 29.5 h post-release, and from then on, every subsequent 24 h. Birds that homed successfully were thus considered to have returned to their respective territory within 5, 30, 54, ..., or 256 h post-release. We set the last visit to 10 days after translocation because such a delay was long enough so that an absence was unlikely to be caused by a lack of navigational ability; short enough so that a presence would not go by undetected, as the birds would still be expected to exhibit territorial behavior independently of their breeding outcome; and because a previous homing experiment (Bélisle et al. 2001) showed that birds are unlikely to return to their territories if they did not do so within 10 days (M. Bélisle unpublished data).
Landscape characterizationForest cover was estimated dichotomously along the trajectories connecting capture and release points using ArcView 3.1 Geographic Information System (ESRI 1996). We coded all mature forest, including burned forest with standing snags, as 1, and all nonforest, including rivers, open bogs, shrub meadows, and anthropogenic features as 0. Because we could not be certain how precisely birds were able to navigate, we calculated forest cover for the 5-m grid cells depicting this capture-to-release line, as well as for the grid cells contained in rectangles extending 100, 200, and 400 m from both the line and from its start and end points. These values were then considered as alternative explanatory variables in our analysis of homing time.
Statistical analysesWe assessed the influence of explanatory variables on the homing time of translocated birds with a Cox regression that was computed using the PHREG procedure in SAS 8.01 (Allison 1995; see Fox [1993] as well as Moya-Larano and Wise [2000] for a discussion of Cox regression analysis applied to ecological problems). Explanatory variables included translocation distance, percent forest cover, treatment (along vs. across barriers), bird species (coded as two dummy variables), and the biologically anticipated interaction between treatment and species (the acknowledgement that species may differ by migratory habits in their response to the treatments). We also included a time-dependent treatment effect by multiplying the treatment variable with homing time because the birds’ motivation to home is likely to decrease with time (see Allison 1995:155-157). We did not consider interactions that included either translocation distance or forest cover, as the magnitudes of these two variables were quite similar within and among all treatment by species combinations (Table 1). We expected a longer homing time for birds translocated across the valley, and accordingly, employed a one-tailed test to quantify the effect of landscape configuration. A one-tailed test was also used to verify that the treatment effect decreased with time. All other tests were two-tailed. We assessed the significance of the Cox regression model and its specific parameters with the likelihood-ratio statistic based on maximum partial likelihood estimates approximated by Efron’s method (see Allison 1995:127-137). Birds that did not return to their territories within 256 h were treated as singly Type I, right-censored data in the Cox regression (sensu Allison 1995). In the context of our experiment, right-censored data are those observations for which we only know that the homing time is greater than a given value, namely our maximum of 256 h. These observations are further considered as being of a single censoring category and Type I when all of the observations are subject to the same censoring time (i.e., 256 h), and when the censoring time is under the control of the researcher (i.e., fixed), respectively. To consider the censoring time as fixed, we had to assume that translocated birds did not experience any significant mortality during the 256 h following translocation. Model R2 was calculated as 1 - exp(G/n), where G is the likelihood-ratio chi-square statistic, andn is the sample size (see Allison 1995:247-249). We set the Type I error rate at 0.05.
Although there was some variation in both forest cover and relocation distance in our homing trials, we are confident that this variation had no systematic effect on the results of our experiment. First, forest cover along the potential routes taken by the homing birds was strongly correlated across spatial scales (Pearson’s product-moment correlation, 100 m vs. 200 m: r = 0.93; 100 m vs. 400 m: r = 0.80; 200 m vs. 400 m: r = 0.94). Given this and the fact that forest birds may sometimes travel more than 250 m under forest cover to avoid crossing an open area (M. Bélisle, unpublished data), we decided to treat forest cover measured within 400 m from all points of the line connecting the capture and release sites as the covariable reflecting landscape composition. Second, translocation distance and forest cover did not vary much among treatment by species combinations (Table 1).
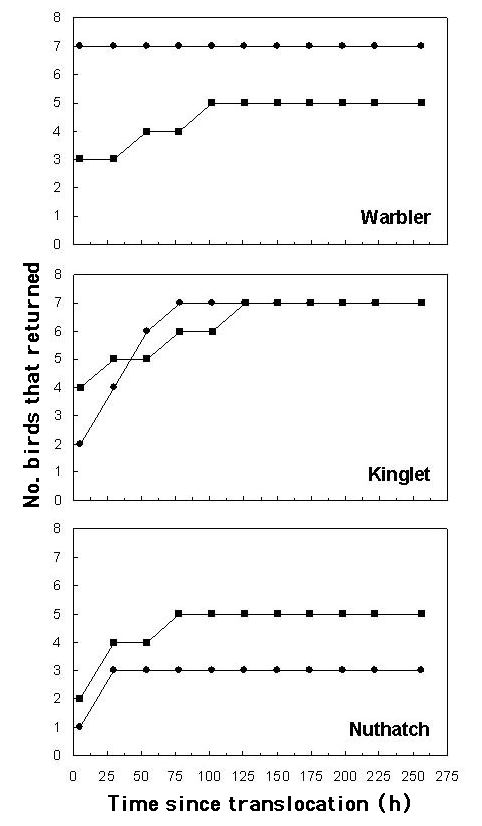
Overall, birds took longer to return to their territories when they were translocated across, rather than parallel to, the barriers, but variation among species belies the generality of this result. As we had predicted, Yellow-rumped Warblers took more time to return to their territories when forced to cross open areas. Indeed, the number of homing events per hour (i.e., hazard in the context of Cox regression; see Allison 1995:15-19) for birds translocated across the valley was estimated to be only 13% of the number of homing events for birds translocated parallel to the valley. (Note that this ratio of hazards, or risk ratio, is obtained by raising e to the value of the parameter estimating the influence of the variable of interest, i.e., e-2.02; see Allison 1995:117.) However, the orientation of barriers did not affect the homing time of Golden-crowned Kinglets. For them, the negative effect of treatment (across) and the positive treatment by species interaction (i.e., across and kinglet) regression coefficients effectively cancelled one another. Contrary to our general expectation, Red-breasted Nuthatches returned faster to their territories when they were translocated across the valley. Also notable was the fact that nuthatches exhibited the poorest return success (57% vs.86% for warblers and 100% for kinglets) and the shortest apparent giving up times, as none returned later than three days post-translocation, whereas a warbler and a kinglet appeared in their territories at four and five days, respectively. Contrary to expectations, the effect of landscape configuration (parallel vs. perpendicular barriers) on homing time did not decrease over time (Table 2, Fig. 2). As expected, forest cover had no bearing on homing time (Table 2), but there was a tendency for birds to take longer to return to their territories when translocated over larger distances (Table 2). Inclusion of a distance by treatment interaction did not improve our regression model (G = 0.5, df = 1, P = 0.46), indicating that translocation distance did not affect the response of birds to translocation treatment.
Table 2. Influence of landscape configuration on the homing time of translocated territorial, mated male Red-breasted Nuthatches, Golden-crowned Kinglets, and Yellow-rumped Warblers (reference category) determined by Cox regression (G = 18.9, df = 8, P = 0.02, R2 = 0.36; n = 42; seven individuals per treatment per species; eight censored cases). All tests are two-tailed except for treatment and treatment x time, where birds translocated along potential barriers serve as the reference category.
a Forest cover was estimated in 5-m grid cells along the capture-to-release line as well as in the grid cells contained in rectangles radiating 400 m from both the line and from its start and end points. b The influence of species and of the interaction between treatment and species, which are both determined by dummy variables, were tested as a whole (by comparing the deviance of the reduced model to the deviance of the full model) before testing the specific effect of their inherent variables. Note that the Yellow-rumped Warbler and the “along barrier” treatment served as reference categories. It follows, for instance, that “across and kinglet” refers to the influence of the bird being a kinglet translocated across the barriers as compared to a warbler translocated along the barriers. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Fig. 2. Homing success of translocated territorial, mated male Yellow-rumped Warblers Dendroica coronata, Golden-crowned Kinglets Regulus satrapa, and Red-breasted Nuthatches Sitta canadensis (n = 42; 7 individuals per treatment per species). Circles and squares depict birds translocated, respectively, along and across potential barriers to movement in the Bow River Valley bottom, Banff National Park, Canada, in 2000.
|
When we consider our results all together, our experiment shows that small open areas running parallel to one another can impede landscape-scale movements of some forest birds, just as a manager’s intuition or a theorist’s understanding of landscape anisotropy would have predicted. We also can be confident that the cumulative barrier effect that we report is attributable to landscape configuration alone because we were able to avoid the confounding effects of landscape composition by standardizing the amount of forest cover in the landscape sections where birds were translocated. Our design and results thus support the prediction that the functional connectivity of landscapes can be an anisotropic attribute requiring that the axes and even the direction of potential animal movement be acknowledged if we are to predict how habitat loss and fragmentation affect movement behavior and, ultimately, dispersal.
All that said, it is also clear that it would be misleading and even erroneous to generalize this result across species, even within the guild of forest birds. Although Yellow-rumped Warblers were strongly impeded by having to cross the parallel barriers, Golden-crowned Kinglets experienced similar homing time regardless of whether individuals had to move along or across these open areas. Rather unexpectedly, Red-breasted Nuthatches appeared to be facilitated by the presence of gaps perpendicular to their direction of travel. The differences among species responses in our experiment are particularly surprising because all three species were previously shown to have a lower propensity to cross open areas with widths as small as 50 m, relative to their willingness to move comparable distances under forest cover (Desrochers and Hannon 1997). Clearly, there is strong species specificity in the effect of landscape configuration on homing time, suggesting that the same may be true of dispersal. There are some potential explanations for the species differences in our study that may also pertain to the dispersal context. Perhaps we did not find a clear treatment effect for kinglets because we measured homing time too crudely. Yet it is difficult to believe that biologically important effects in the context of landscape-scale dispersal could be masked by our diel schedule. It is more likely that kinglets were genuinely unaffected by these barriers at this scale (2–3 km) of movement. Understanding the response of nuthatches, whose movement appeared to be actually enhanced by the barriers, is more difficult. One possibility is that nuthatches, as facultative year-round residents, were prospecting for better territories upon their return paths and simply lost the motivation to return home when a superior territory was encountered. We noticed at least two translocated individuals that appeared to have taken up residence in between their capture and release points. Another possibility is that nuthatches translocated across the valley were able to use the highway as a landmark that clearly indicated the direction in which their territories were located, whereas nuthatches translocated parallel to the valley did not benefit from such an informational advantage (Cannicci et al. 2000). Notwithstanding these plausible explanations, we hesitate to interpret the counterintuitive response of the nuthatches in the context of movement constraints, simply because they could result from a lack of motivation or navigational ability on the part of the individuals.
Suffice it to say that our unexpected species differences have some implications for future empirical work of this sort. First, we must know something about a species’ homing ability before we can interpret the results of homing experiments in the context of dispersal. Second, our results warn against directly transposing the findings of short-term, small-scale experiments on movement behavior to the landscape scale. This is true even though these experiments may have considerable potential to predict species’ occupancy patterns in the landscape (Desrochers et al. 1999, Jonsen and Taylor 2000). A third implication stemming from these species differences is that more precise information about the return trajectories is needed. Were researchers able to follow birds as they returned, they would be able to better measure the effects of individual landscape features on both the timing and success with which birds move through fragmented habitats.
Assuming that our results are not artifacts of differences in homing abilities or responses to particular features, they may shed new light on assumptions about the effects of habitat loss and fragmentation on forest birds more generally. It is a common, but largely untested, belief that open areas should impede the movements of migratory forest bird species to a lesser extent than the movements of resident species (Hunter 1992, Wiens 1994). This assumption stems from the fact that Neotropical migrants cross whole continents and large bodies of water annually; it thus seems inconceivable that they could be impeded by single linear features like highways. Our results, however, caution against blindly using the migratory status of a species as an index of its sensitivity to movement constraints stemming from either landscape composition or configuration. Indeed, warblers (long-distance migrants) returned to their territories less rapidly than kinglets (short-distance migrants). Likewise, in a similar homing experiment conducted within an agricultural landscape by Bélisle et al. (2001), no difference in homing time was observed among two long-distance migrants, namely the Black-throated Blue Warbler (Dendroica caerulescens) and the Ovenbird (Seiurus aurocapillus), and the resident Black-capped Chickadee (Poecile atricapillus). Movement in the opposite direction or lack of variation among species suggests that birds are susceptible to being disrupted in their dispersal movements by habitat loss and fragmentation independently of their migratory status. This generalization may not persist during migration, when birds would presumably have greater motivation to move directionally and thereby show less sensitivity to barrier effects during their daily movements. The probable importance of motivational context to the movement of birds exemplifies the need to better acknowledge a variety of behavioral attributes in conservation questions (Lima and Zollner 1996, Caro 1998, Sutherland 1998). Certainly it is clear that data will have to be collected on a wider array of species and in different seasons before we can properly assess the importance of migratory status, either within or among species, as a predictor of a species’ sensitivity to movement constraints imposed by habitat loss and fragmentation.
Perhaps the most important conclusion of our experiment is that small gaps in forest cover that momentarily thwart the movements of forest birds (Desrochers and Hannon 1997, St. Clair et al. 1998) can add up to cause a significant cumulative barrier effect at the landscape scale, at least for some species. When these gaps contain barriers like high-volume highways, with their associated noise deterrents (Reijnen et al. 1995), the effect is likely to be exacerbated. Such barrier effects, combined with the propensity to follow forest boundaries that is exhibited by some forest birds (Desrochers et al. 1999, Desrochers and Fortin 2000), have considerable potential to polarize the movement rates of dispersing individuals. Polarized emigration and immigration rates could have important influences on population structure and dynamics, particularly if dispersing individuals are channeled in such a way that some key habitat patches for the persistence of a population network are repeatedly, yet passively, avoided (Hanski and Ovaskainen 2000). For this reason, developing methods that will allow us to measure functional connectivity directionally along different axes of landscape sections will certainly be of interest to both fundamental and applied ecologists. An interesting avenue toward this aim is the implementation of empirical, gap-crossing decision rules of forest birds into “realistic” percolation or graph models (Keitt et al. 1997, Urban and Keitt 2001) or spatially explicit, individual-based models (Gustafson and Gardner 1996, Schippers et al. 1996, Schumaker 1996, Brooker et al. 1999, Wiegand et al. 1999). Such models could employ Monte Carlo simulations to assess the likelihood of cyber-birds to cross real landscape sections along various axes, and in different directions along these axes. With such reality-based models, we could hope to better predict the effects of disruptions to habitat, ultimately also mitigating and even adaptively planning these in our increasingly human-altered landscapes.
Responses to this article are invited. If accepted for publication, your response will be hyperlinked to the article. To submit a comment, follow this link. To read comments already accepted, follow this link.
We thank E. Kloppers, A. Roe, and C. Thiessen for their generous help in the field. D. Zell (Parks Canada) provided GIS maps and ortho-corrected aerial photos of the study area, and J. Whittington calculated the forest cover information. Parks Canada and T. Clevenger provided important assistance throughout the study, and A. Desrochers, L. Imbeau, and M. J. Mazerolle provided insightful comments on previous drafts of this paper. This work was supported by a NSERC (Natural Sciences and Engineering Research Council of Canada) research grant to C. C. St. Clair, the Office of the Vice-President (Research), University of Alberta, and a Challenge Grant in Biodiversity from the Alberta Conservation Association to M. Bélisle.
Allison, P. D. 1995. Survival analysis using the SAS system. SAS Institute, Cary, North Carolina, USA.
Anders, A. D., J. Faaborg, and F. R. Thompson, III. 1998. Postfledging dispersal, habitat use, and home-range size of juvenile Wood Thrushes. Auk 115:349-358.
Bélisle, M., A. Desrochers, and M.-J. Fortin. 2001. Influence of forest cover on the movements of forest birds: a homing experiment. Ecology 82:1893-1904.
Bender, D. J., T. A. Contreras, and L. Fahrig. 1998. Habitat loss and population decline: a meta-analysis of the patch size effect. Ecology 79:517-533.
Brooker, L., M. Brooker, and P. Cale. 1999. Animal dispersal in fragmented habitat: measuring habitat connectivity, corridor use, and dispersal mortality.Conservation Ecology [Online] 3(1):4. URL: http://www.consecol.org/vol3/iss1/art4/.
Cannicci, S., C. Barelli, and M. Vannini. 2000. Homing in the swimming crab Thalamita crenata: a mechanism based on underwater landmark memory. Animal Behaviour 60:203-210.
Caro, T., editor. 1998. Behavioral ecology and conservation ecology. Oxford University Press, Oxford, UK.
Clevenger, A. P., and N. Waltho. 2000. Factors influencing the effectiveness of wildlife underpasses in Banff National Park, Alberta, Canada. Conservation Biology 14:47-55.
Desrochers, A., and M.-J. Fortin. 2000. Understanding avian responses to forest boundaries: a case study with chickadee winter flocks. Oikos 91:376-384.
Desrochers, A., and S. J. Hannon. 1997. Gap crossing decisions by forest songbirds during the post-fledging period. Conservation Biology 11:1204-1210.
Desrochers, A., S. J. Hannon, M. Bélisle, and C. C. St. Clair. 1999. Movement of songbirds in fragmented forests: Can we "scale up" from behaviour to explain occupancy patterns in the landscape? Pages 2447-2464 in N. Adams and R. Slotow, editors. International Ornithological Congress 22:2447-2464.
ESRI. 1996. ArcView GIS version 3.11. Environmental Systems Research Institute, Redlands, California, USA.
Fahrig, L. 1997. Relative effects of habitat loss and fragmentation on population extinction. Journal of Wildlife Management 61:603-610.
Forman, R. T. T., and L.E. Alexander. 1998. Roads and their major ecological effects. Annual Review of Ecology and Systematics 29:207-231.
Fox, G. A. 1993. Failure-time analysis: emergence, flowering, survivorship, and other waiting times. Pages 253-289 in S. M. Scheiner and J. Gurevitch, editors. Design and analysis of ecological experiments. Chapman and Hall, New York, New York, USA.
Ghalambor, C. K., and T. E. Martin. 1999. Red-breasted nuthatch, Sitta canadensis. In A. F. Poole, P. Stettenheim, and F. B. Gill, editors. The birds of North America. Number 459. American Ornithologists' Union and Academy of Natural Sciences, Washington, D.C., and Philadelphia, Pennsylvania, USA.
Grubb, T. C., and P. F. Doherty. 1999. On home-range gap-crossing. Auk 116:618-628.
Gustafson, E. J., and R. H. Gardner. 1996. The effect of landscape heterogeneity on the probability of patch colonization. Ecology 77:94-107.
Haines-Young, R., and M. Chopping. 1996. Quantifying landscape structure: a review of landscape indices and their application to forested landscapes. Progress in Physical Geography20:418-445.
Hargis, C. D., J. A. Bissonette, and J. L. David. 1998. The behavior of landscape metrics commonly used in the study of habitat fragmentation. Landscape Ecology 13:167-186.
Hinsley, S. A. 2000. The costs of multiple patch use by birds. Landscape Ecology 15:765-775.
Hunt, P. D., and D. J. Flaspohler. 1998. Yellow-rumped Warbler, Dendroica coronata. In A. F. Poole, P. Stettenheim, and F. B. Gill, editors. The birds of North America. Number 376. American Ornithologists' Union and Academy of Natural Sciences, Washington, D.C., and Philadelphia, Pennsylvania, USA.
Hunter, M. L. J. 1992. Paleoecology, landscape ecology, and conservation of neotropical migrant passerines in boreal forests. Pages 511-523in J. M. Hagan, III and D. W. Johnston, editors. Ecology and conservation of neotropical migrant landbirds. Smithsonian Institution Press, Washington D. C., USA.
Ingold, J. L., and R. Galati. 1997 Golden-crowned Kinglet, Regulus satrapa.In A. F. Poole, P. Stettenheim, and F. B. Gill, editors. The birds of North America. Number 301. American Ornithologists' Union and Academy of Natural Sciences, Washington, D.C., and Philadelphia, Pennsylvania, USA.
Jonsen, I., and P. D. Taylor. 2000. Calopteryx damselfly dispersions arising from multiscale responses to landscape structure. Conservation Ecology [online] 4(2): 4. URL: http://www.consecol.org/vol4/iss2/art4/.
Keitt, T. H., D. L. Urban, and B. T. Milne. 1997. Detecting critical scales in fragmented landscapes. Conservation Ecology [online] 1(1):4. URL: http://www.consecol.org/vol1/iss1/art4/.
Lima, S. L., and P. A. Zollner. 1996. Towards a behavioral ecology of ecological landscapes. Trends in Ecology and Evolution 11:131-135.
Machtans, C. S., M.-A. Villard, and S. J. Hannon. 1996. Use of riparian buffer strips as movement corridors by forest birds. Conservation Biology 10:1366-1379.
Matthysen, E., F. Adriaensen, and A. A. Dhondt. 1995. Dispersal distances of nuthatches,Sitta europaea, in a highly fragmented forest habitat. Oikos 72:375-381.
McGarigal, K., and B. J. Marks. 1995. FRAGSTATS: spatial pattern analysis program for quantifying landscape structure. U.S. Forest Service General Technique Report PNW 351.
Moya-Larano, J., and D. H. Wise. 2000. Survival regression analysis: a powerful tool for evaluating fighting and assessment. Animal Behaviour 60:307-313.
Page, R., S. Bayley, J. D. Cook, J. E. Green, and J. R. B. Ritchie. 1996. Banff-Bow Valley: At the Crossroads. Summary report of the Banff-Bow Valley Task Force. Department of Canadian Heritage, Ottawa, Ontario, Canada.
Paradis, E., S. Baillie, W. Sutherland, and R. Gregory. 1998. Patterns of natal and breeding dispersal in birds. Journal of Animal Ecology 67:518 - 536.
Rail, J.-F., M. Darveau, A. Desrochers, and J. Huot. 1997. Territorial responses of boreal forest birds to habitat gaps. Condor 99:976-980.
Reijnen, R., R. Foppen, C. ter Braak, and J. Thissen. 1995. The effects of car traffic on breeding bird populations in woodland. III. Reduction of density in relation to the proximity of main roads. Journal of Applied Ecology 32:187-202.
Schippers, P., J. Verboom, J. P. Knaapen, and R. C. van Apeldoorn. 1996. Dispersal and habitat connectivity in complex hetrogeneous landscapes: an analysis with a GIS-based random walk model. Ecography 19:97-106.
Schumaker, N. H. 1996. Using landscape indices to predict habitat connectivity. Ecology 77:1210-1225.
St. Clair, C. C., M. Bélisle, A. Desrochers, and S. J. Hannon. 1998. Winter response of forest birds to habitat corridors and gaps. Conservation Ecology [online]2(2):13. URL: http://www.consecol.org/vol2/iss2/art13/.
Sutherland, W. J. 1998. The importance of behavioral studies in conservation biology. Animal Behaviour 56:801-809.
Tischendorf, L. 2001. Can landscape indices predict ecological processes consistently?Landscape Ecology 16:235-254.
Tischendorf, L., and L. Fahrig. 2000. On the usage and measurement of landscape connectivity. Oikos 90: 7-19.
Trombulak, S. C., and C. A. Frissell. 2000. Review of ecological effects of roads on terrestrial and aquatic communities. Conservation Biology 14:18-30.
Turchin, P. 1998. Quantitative analysis of movement. Sinauer Associates, Sunderland, Massachusetts, USA.
Urban, D., and T. Keitt. 2001. Landscape connectivity: a graph-theoretic perspective.Ecology 82:1205-1218.
Vega Rivera, J. H., J. H. Rappole, W. J. McShea, and C. A. Haas. 1998. Wood Thrush postfledging movements and habitat use in northern Virginia. Condor 100:69-78.
van Langevelde, F. 2000. Scale of habitat connectivity and colonization in fragmented nuthatch populations. Ecography 23:614-622.
Verboom, J., A. Schotman, P. Opdam, and H. Metz. 1991. European nuthatch metapopulations in a fragmented agricultural landscape. Oikos 61:149-156.
Verhulst, S., C. M. Perrins, and R. Riddington. 1997. Natal dispersal of Great Tits in a patchy environment. Ecology 78:864-872.
Villard, M.-A., G. Merriam, and B. A. Maurer. 1995. Dynamics in subdivided populations of neotropical migratory birds in a fragmented temperate forest. Ecology 76:27-40.
Villard, M.-A., M. K. Trzcinski, and G. Merriam. 1999. Fragmentation effects on forest birds: relative influence of woodland cover and configuration on landscape occupancy. Conservation Biology 13:774-783.
Wiegand, T., K. A. Moloney, J. Naves, and F. Knauer. 1999. Finding the missing link between landscape structure and population dynamics: a spatially explicit perspective. American Naturalist 154:605-627.
Wiens, J. A. 1994. Habitat fragmentation: islandv. landscape perspectives on bird conservation. Ibis 137:S97-S104.
Wilcox, L, B. Robinson, and A. Harvey. 1998. A sense of place: An atlas of issues, attitudes and resources in the Yellowstone to Yukon Ecoregion. Yellowstone to Yukon Conservation Initiative, Canmore, Alberta, Canada.
Address of Correspondent:
Marc Bélisle
Present address:
Metapopulation Research Group, Department of Ecology and Systematics
University of Helsinki, P.O. Box 17, Arkadiankatu 7
FIN-0014, Helsinki, Finland
Phone: +358-9-191-28778
belisle_marc@hotmail.com
![]()