|
|
|
Copyright © 2001 by the author(s). Published here under license by The Resilience Alliance.
The following is the established format for referencing this article:
Elmqvist, T., M. Wall, A. L. Berggren, L. Blix, Å. Fritioff, and U. Rinman. 2001. Tropical forest reorganization after cyclone and fire disturbance in Samoa: remnant trees as biological legacies. Conservation Ecology 5(2): 10. [online] URL: http://www.consecol.org/vol5/iss2/art10/
A version of this article in which text, figures, tables, and appendices are separate files may be found by following this link.
Report Tropical Forest Reorganization after Cyclone and Fire Disturbance in Samoa: Remnant Trees as Biological Legacies Thomas Elmqvist, Maria Wall, Anna-Lena Berggren, Lisa Blix, Åsa Fritioff, and Ulrika Rinman
Stockholm University
- Abstract
- Introduction
- Methods
- Results
- Discussion
- Conclusion
- Responses to this Article
- Acknowledgments
- Literature Cited
In disturbed rain forests, large, living remnant trees may be of significant importance for postdisturbance reorganization either directly, by producing large quantities of seeds, or indirectly, by attracting vertebrate seed dispersers. In addition, remnant trees may also be important in providing a favorable microhabitat for seedlings of late-successional species. This study focused on the role of large remnant trees (> 40 cm dbh) in patterns of regeneration after cyclone and fire damage in the Tafua and Falealupo Rain Forest Preserves, Savaií, Samoa. At Tafua, 10 large trees at each of two sites (one site burned in 1990) were investigated with regard to numbers of species and densities of plants from three different size classes at different distances from remnant trees. At the burned site, both species richness and the densities of plants < 1cm dbh were significantly higher inside the canopies of remnant trees than outside of them. At the unburned site, no or only marginally significant differences were observed. At Falealupo, two burned sites (burned in 1993 and 1998) were investigated using seed traps. At both sites, the seed rain from vertebrate dispersers was disproportionally higher under the canopies of remnant trees than in outside areas. No differences in soil characteristics were found when comparing samples taken from inside and outside canopies. Our results are congruent with the prediction that large remnant trees surviving in severely disturbed rain-forest areas represent biological legacies and serve as nuclei for reorganization. Based on this study and our previous work, we suggest that three factors represent essential components of the spatial resilience of tropical forest ecosystems and should be targeted for active management in tropical forests exposed to large-scale disturbances, particularly fire: remnant trees, refugia, and vertebrate dispersers.
KEY WORDS: biological legacies, cyclone, fire, rain forest, remnant trees, reorganization, vertebrate dispersal.
Published: December 20, 2001
Disturbances are integral features of ecosystems, and most ecosystems are subject to several disturbance regimes that take place at different temporal and spatial scales (Holling et al. 1995, Turner et al. 1998). Recent studies on how biotic communities reorganize and re-establish after large-scale disturbances point to the common pattern of nucleation, i.e., the spread of organisms from numerous foci rather than their establishment inward from the margins (Franklin and MacMahon 2000, Franklin et al. 2000). Such foci have been labeled "biological legacies," and may consist of living organisms, organically derived structures, or organically generated spatial patterns (Foster et al. 1998, Franklin et al. 2000). Living remnant trees in disturbed tropical forests represent one example of such biological legacies, and may have both direct and indirect effects on the rate and type of forest regeneration. Large trees may contribute directly to regeneration by producing large quantities of seeds. Indirectly, they may attract seed dispersers such as birds and bats due to the presence of fleshy fruits, nectar-producing flowers, or leaves; they may also provide facilities such as protection and rest (Guevara et al. 1986, Wunderle 1997). In fragmented tropical forests and in tropical pastures, isolated remnant trees have often been found to be important foci for regeneration (Guevara et al. 1986, Guevara et al. 1992, Wunderle 1997, Toh et al. 1999), because the birds or bats that alight in them defecate or regurgitate seeds (Guevara et al. 1986, Guevara and Laborde 1993, Robinson and Handel 1993). The presence of remnant trees may also facilitate establishment, because the conditions prevailing under the tree canopies usually represent favorable microclimatic or nutrient conditions for the seedlings of late-successional plants (Guevara et al. 1986, Belsky et al. 1989, Guevara et al. 1992).
The islands of Samoa were struck by two severe cyclones, Ofa in 1990 and Val in 1991, with wind speeds in excess of 200 km/h (Pandaram 1992, Elmqvist et al. 1994). Afterward, parts of the Falealupo and Tafua Rain Forest Preserves on the island of Savaií were also ravaged by forest fires. The cyclones and the fires caused extensive changes to the forests. At Tafua, the density of trees > 5 cm dbh decreased from 476/ha prior to the cyclones to 225/ha after Val, and to fewer than 43/ha in the burned portion of the forest. The mean canopy cover decreased from nearly 100% before Ofa to 27% in the unburned and 12% in the burned forest after Val (Elmqvist et al. 1994). A few large living trees were still present even in the most intensively burned areas (Elmqvist et al. 1994, Hjärpe et al. 2001).
Animal seed dispersers are generally thought to be of great importance in the regeneration process after catastrophic destruction of tropical forests (e.g., Wunderle 1997). According to Cox et al. (1995), Rainey et al. (1995), Pierson et al. (1996), and Banack (1998), most of the dominant forest canopy and subcanopy trees in Samoa produce fleshy fruits that are consumed by two species of flying foxes (Pteropus spp. and Pteropodidae) and several species of fruit pigeons (e.g., Ducula pacifica and Ptilinopus perousii) and starlings (Sturnus spp.). The purpose of this study was to investigate the role of living remnant trees in attracting seed dispersers and shaping the form and rate of regeneration in areas that had been severely disturbed by cyclones and fire. This seemed to be of particular relevance because Hjärpe et al. (2001), in their study of the burned area at Tafua, concluded that colonization and regeneration were limited by seed dispersal.
To study the role of dispersal and remnant trees in the reorganization phase after disturbance, we addressed the following questions:
- 1) Are the diversity and density of recently established vertebrate-dispersed species higher closer to the remnant trees or farther away?
- 2) Is there a higher multispecies seed rain under the canopies of remnant trees or outside the canopies?
- 3) Are the microclimatic and nutrient conditions for seedling establishment more favorable closer to remnant trees or farther away?
Two sites in the Tafua Peninsula Rain Forest Preserve were selected for the first study, and two sites in the Falealupo Rain Forest Preserve, Savaií, Samoa (13–15° S, 168–173° W), were chosen for the second (Cox and Elmqvist 1991). Rainfall in these areas is approximately 2500 mm/yr with a precipitation peak in October–March. The mean monthly temperature is 26°C. Most of the areas constitute low-relief lava plains < 30 m above sea level. The Tafua forest (~ 5000 ha) is the largest remaining block of continuous lowland rain forest in Samoa. Before the disturbance, the forest was dominated by Pometia pinnata (Sapindaceae); other important species included Dysoxylum maota (Meliaceae), Garuga floribunda (Burseraceae), Planchonella torricellensis (Sapotaceae), and Syzygium inophylloides (Myrtaceae) prior to the cyclones (Whistler 1992, Elmqvist et al. 1994). The first site at Tafua was located on the slope of a small volcanic crater, and the other site was approximately 1 km from the crater. These sites correspond to sites used in earlier studies (Elmqvist et al. 1994, Hjärpe et al. 2001) and were surveyed in both 1992 and 1996. The second site burned intensively in February 1990 (Elmqvist et al. 1994). The density of large remnant trees (> 40 cm dbh) was estimated at 10/ha at the burned site (n = 5, plot size = 400 m2) and 42/ha (n = 14, plot size = 400 m2) at the crater site. The present study was conducted from March through June of 1997.
At Falealupo, the two study sites were located within a burned portion (approximately 100 ha) of the 3000-ha forest preserve (Cox and Elmqvist 1991). The burned area was approximately 3 km from the main road in the part of the reserve that contained the largest contiguous tract of closed-canopy forest prior to the cyclone (Pierson et al. 1996). Two recent fires occurred there, one in August 1993 (referred to in our study as the 1993 site) and in part of this area again in July–August 1998 (referred to as the 1998 site). The fires resulted in more than 90% mortality of canopy trees at the 1993 site and more than 98% at the 1998 site (T. Elmqvist, personal observation). Prior to the cyclones and fires, the forest was dominated by Garuga floribunda, Pometia pinnata, and Syzygium spp. (T. Elmqvist, personal observation; see also Whistler 1992). At the 1993 site, a few large trees > 40 cm dbh were still alive; the predominant species was G. floribunda, but there were also a few Terminalia catappa, Ficus spp., Planchonella torricellensis, and Syzygium inophylloides. At the 1998 site, only a few G. floribunda remained. This study was conducted during October and November of 1998.
At both sites, open sun-exposed ground was covered by Mikania micrantha and, to a lesser extent, by Passiflora foetida. Other common and typical species in the open spaces between remnant trees were Macaranga harveyana, Carica papaya, Morinda citrifolia, and Pipturus argenteus. Under the canopies of the remnant trees, the number of species was higher, and more primary forest trees were represented, including P. pinnata, Syzygium spp., Dysoxylum samoense, Ficus spp., G. floribunda, and Diospyros samoensis.
Study 1: TafuaAt each of the two sites, 10 remnant trees > 40 cm dbh were haphazardly chosen, with enough distance between them so that transects would not overlap. For each tree, the diameter and height were determined (Table 1) using a diameter tape and a Silva Clinomaster height meter with two scales. Because the crater site was narrow, the trees chosen there were surrounded by enough open area to permit transects without having plantation land or other transects cross the sample site. Transects were put out in the four cardinal compass points for every tree, to make sure that eventual differences in composition were not due to differences in direction. Sample plots with areas of 4 m2 were put out with their centers at distances of 5, 10, 15, and 20 m, respectively, from each tree in each of the four transects. The sample plots closest to the trees under the canopy had areas of 6 m2. This design was chosen to compensate in part for the larger total area sampled outside the canopies (640 vs. 240 m2 at each site). The plots at a distance of 5 m were typically located at the margin of or outside the canopy. Within each sample plot, only woody plants were examined; ferns, herbs, and vines were excluded. The shrub Aidia cochinchinensis (Rubiaceae), a fairly common species at the crater site but very rare in the burned area, was also excluded, because it tends to form large clones whose genets are difficult to separate and because it is propagated mainly by cloning rather than by vertebrate dispersal.
|
Table 1. Characteristics of remnant trees sampled in Study 1 at the Tafua Rain Forest Preserve and Study 2 at the Falealupo Rain Forest Preserve, Samoa.
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
The numbers of individuals in each species were counted, and every plant was sorted into one of three size classes: < 1 cm, 1 > 5 cm, and 5 > 20 cm dbh. Species richness (number of species per square meter) and plant density (number of individuals per square meter) were calculated for each size class at each site. Species richness values and plant densities were also calculated for every size class at all sample plot distances at both sites. In the sample plots closest to remnant trees, all records of plants of the same species as the tree itself were excluded from the analyses to obtain a more conservative estimate of dispersal. When testing for differences in density or species richness inside and outside canopies, a mean for each tree and size class was calculated based on sample plots inside the canopy and on the pooled sample plots from outside the canopy; these values were used in further statistical analyses. More detailed analyses were performed on the smallest size class, because those plants were the most likely to have established after the cyclone and fire.
One group of species, Syzygium spp., which is known to be highly preferred by flying foxes (Banack 1998), was analyzed separately based on all records of plants < 5 cm dbh at all sample plot distances at both sites.
Statistical analyses
Analyses of variance were used to compare mean values when group variances were the same (Levene's test). Groups with uneven sample sizes and unequal variances were compared using Mann-Whitney U-tests.
Study 2: FalealupoA total of 13 remnant trees was identified, seven at the 1993 site and six at the 1998 site. Two sets of two seed traps each were set out at each tree. The seed traps were made of fiber sheets measuring 1.5 x 2 m. The surface of the sheets was coarse and prevented seeds from being washed away when it rained. The fiber also allowed a certain amount of rainwater to permeate through. The first two traps were placed immediately under the canopy, near the trunk, in randomly selected directions. The second two were placed in randomly selected directions at a distance of 20 m from the tree trunk.
At the 1993 site, the seed traps were left out for 26 days and checked 17 times. At the 1998 site, the traps were left out only for 13 days, during which they were checked seven times. Normally, the traps were surveyed between 0600 and 1100, but on one or two occasions the check was carried out in the afternoon. The collected seeds were counted, and the species determined. For the seed traps closest to the trees, all records of seeds of the same species as the tree itself were excluded from the analyses.
Soil
Soil samples were taken within 50 cm of each seed trap. Samples were dug to a depth of about 20 cm in an area approximately 50 cm2. The soil samples were placed in plastic bags, which were left open to allow the soil to air-dry. The samples were transported to the Department of Soil Science at the University of the South Pacific in Alafua, Samoa. At the soil laboratory, the samples were analyzed in terms of moisture factor, pH (water), total nitrogen, NH4OAc-extractable calcium, magnesium, potassium, and Truog phosphorus.
Shading and vegetation cover
Two sample squares, each measuring 0.25 m2, were randomly placed near each seed trap. The percentages of the ground exposed to direct sunlight or covered with vegetation, dead wood, or rocks were estimated and assigned to one of the following categories: 0–20%, 20–40%, 40–60%, 60–80%, or 80–100% of the sample square.
Statistical analyses
Data on numbers of seeds and species numbers were log-transformed (log y + 0.1) to improve normality and analyzed using ANOVA. Results of soil samples were analyzed by the Wilcoxon matched-pair sign rank test, with P values adjusted via the Bonferoni method. Proportion values were arcsine-transformed.
As shown in Table 1, the 10 remnant trees included in the study at the burned site were represented by three species: Terminalia catappa, Garuga floribunda, and Pometia pinnata. At the crater site, five species were represented: P. pinnata, G. floribunda, Dysoxylum maota, T. catappa, and Syzygium inophyllides. Their mean dbh was 81.8 cm, and their mean height was 29.2 m. Most of these species produce fleshy fruits, but none were observed to be fruiting at the time of study.
Differences between sites
The total number of woody plants examined in the transects was 2748 at the crater site and 1171 at the burned site. Total species richness was 48 species at the crater site and 41 at the burned site. Canarium vitense, Cryptokaria elegans, Premna serratifolia, Inocarpus fagifer, Planchonella torricilencis, Psychotria insularum, Sterculia adenanthera, Fagrea sp., and Cypholopus stipulatus were found only at the crater site, whereas Alphitonia sp., Castilla elastica, Syzygium savaiense, Carica papaya, and Psidium sp. were found only at the burned site. At the crater and burned sites, respectively, there were 2420 and 786 plants in the first size class (< 1 cm in diameter), 253 and 215 in the second (1 < 5 cm in diameter), and 75 and 170 in the third (5 < 20 cm in diameter). The plants in the smallest size class had significantly higher mean values for both plant density and species richness at the crater site than at the burned site (ANOVA, P < 0.001). In the medium size class, no significant difference in either plant density or species richness was found between the sites (ANOVA, P > 0.05). The plants in the largest size class had significantly higher densities at the burned site (ANOVA, P < 0.001) (see Table 2).
|
Table 2. Species richness, expressed as number of species per square meter, and density, expressed as number of individuals per square meter, in three different size classes of plants at different distances from remnant trees at the burned site (n = 10) and the crater site (n = 10) in the Tafua Rain Forest Preserve, Samoa. Mean is ± 1 SE.
†Test of difference in species richness or plant density comparing samples from inside the canopies with pooled samples from outside the canopies (Mann-Whitney U-test). *** = P < 0.001, **= P < 0.01, * = P ~ 0.05. ‡Test of differences among sites for each size class (ANOVA), *** = P < 0.001. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Differences among plants located inside and outside the canopies
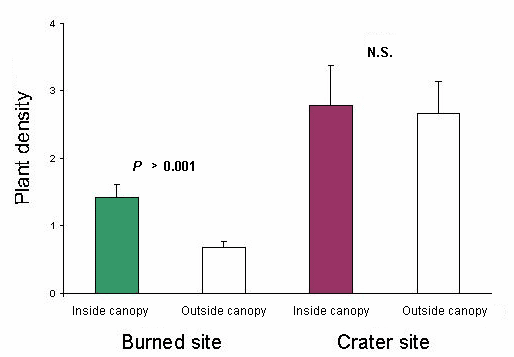
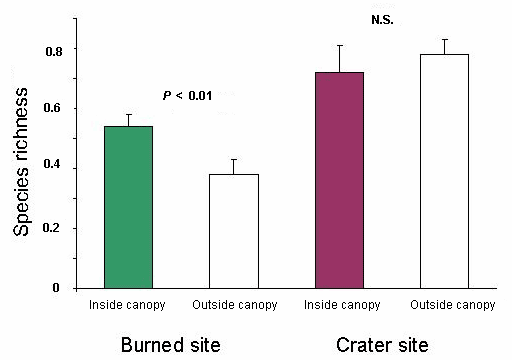
At the burned site and among the plants in the smallest size class, a total of 341 plants belonging to 13 species was recorded inside the canopies (a total sample area of 240 m2), and 435 plants belonging to 26 species were recorded outside the canopies (a total sample area of 640 m2). As seen in Table 2 and Fig. 1, plant density was significantly higher under the canopies than outside of canopy cover (Mann-Whitney, P < 0.001). Also, in the smallest size class, significantly higher species richness (species per square meter) was found in the sample plots closest to the trees than outside the canopies (Mann-Whitney, P < 0.01), as shown in Table 2 and Fig. 2. Among medium-sized and large plants, no difference in plant density inside vs. outside the canopies was found Table 2). Species richness tended to be higher in samples taken outside the canopy for medium-sized plants (Mann-Whitney, P ~ 0.05), whereas no difference was found for large plants (Table 2). Six of the 10 most abundant species in the smallest size class found close to the trees were classified as late-successional canopy species, whereas outside the canopies this relation was five out of 10.
|
Fig. 1. Plant densities expressed as number of individuals per square meter for plants < 1 cm dbh inside and outside canopies of remnant trees at the burned site and the crater site in the Tafua Rain Forest Preserve, Samoa (n = 10). Mean is ± SE.
|
|
Fig. 2. Species richness expressed as number of species per square meter for plants < 1 cm dbh inside and outside canopies of remnant trees at the burned site and the crater site in the Tafua Rain Forest Preserve, Samoa (n = 10). Mean is ± SE.
|
At the crater site and among the plants of the smallest size class, a total of 670 plants belonging to 15 species was recorded inside the canopies (a total sample area of 240 m2), and 1710 plants belonging to 29 species were recorded outside the canopies (a total sample area of 640 m2). There were no significant differences between samples taken from inside and outside the canopies in terms of plant density or species richness in the small size class (Figs. 1 and 2), or in the medium and large size class (Mann-Whitney, all tests, P > 0.05), as shown in (Table 2). For the plants in the smallest size class, there was a pronounced trend toward a peak in both density and species richness at a distance of 5 m (Table 2).
Differences at different sampling distances for Syzygium spp.
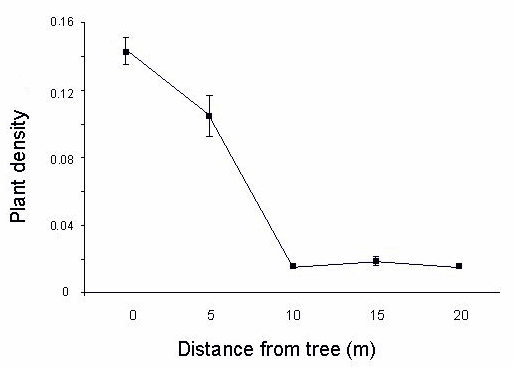
As seen in Fig. 3, individuals of Syzygium spp. < 5 cm dbh at the burned site had significantly higher densities close to remnant trees than outside the canopies (Mann-Whitney, P < 0.05). At the crater site, individuals of Syzygium spp. < 5 cm dbh were much more uncommon, with a mean density of 0.03 ± 0.008/m2 and with no detectable pattern at different distances from trees (Mann-Whitney, P > 0.05).
|
Fig. 3. Density expressed as plants per square meter of Syzygium spp. < 5 cm dbh at different distances from remnant trees at the burned site in theTafua Rain Forest Preserve, Samoa (n = 10).
|
Study 2: Falealupo
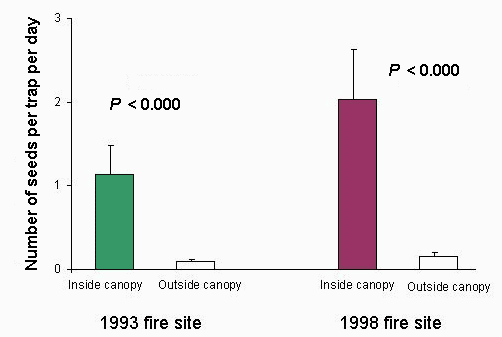
All the trees used in the study were G. floribunda, except for one large strangler fig (Ficus sp.) and one P. torricellensis. At the 1993 site, a total of 728 trap days was included in the study, with 312 trap days at the 1998 site. The total number of seeds dropped into seed traps was 266 inside and 22 outside the canopies at the 1993 site and 170 inside and 13 outside the canopies at the 1998 site. As shown in Fig. 4, this difference was significant at both sites (ANOVA, F1.22 = 21.5, P < 0.000). Similarly, Fig. 5 shows that the species richness of the seed samples was significantly higher under tree canopies than outside of them at both sites (ANOVA, F1.22 = 18.5, P < 0.000). The dominant early-successional, vertebrate-dispersed species found in the seed traps were Macaranga harveyana, Carica papaya, Castillea elastica, and Morinda citrifolia, whereas Bischofia javonica, Dysoxylum samoense, and Ficus spp. were the principal late-successional, vertebrate-dispersed species. Seeds of M. harveyana and C. papaya were the most numerous in the traps (> 80%), followed by Ficus spp. and M. citrifolia. Other species were very poorly represented.
|
Fig. 4. Average number of vertebrate-dispersed seeds dropped in seed traps inside and outside canopies of remnant trees at the sites burned in 1993 (n = 7 trees) and 1998 (n = 6 trees) in the Falealupo Rain Forest Preserve, Samoa. Mean is ± SE. The samples were taken during October and November of 1998.
|
|
Fig. 5. Average number of species of vertebrate-dispersed seeds dropped in seed traps inside and outside canopies of remnant trees at sites burned in 1993 (n = 7 trees) and 1998 (n = 6 trees) in the Falealupo Rain Forest Preserve, Samoa. Mean is ± SE. The samples were taken in October and November of 1998.
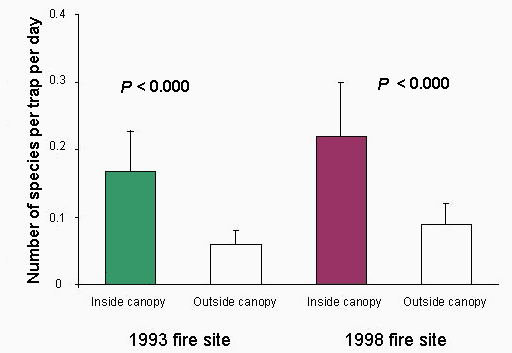
|
Soil analyses
Analyses of moisture factor, pH (water), total nitrogen, NH4OAc-extractable calcium, magnesium, potassium, and Truog phosphorus in soil samples (Table 3) did not show any significant differences when comparing samples from under tree canopies with samples collected from outside of them (Wilcoxon, Bonferoni adjusted p values).
|
Table 3. Results for soil samples taken inside and outside canopies at sites in the Falealupo Rain Forest Preserve, Samoa, that burned in 1993 (n = 7) and 1998 (n = 6). The samples were analyzed for moisture factor (MF) or percentage water content of the soil, the pH of the water contained in the soil, and total nitrogen (N), NH4OAc-extractable calcium (Ca), magnesium (Mg), potassium (K), and Truog phosphorus (P) expressed as grams per kilogram of soil.
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Shading and vegetation cover
At both sites, the proportion of ground area exposed to direct solar radiation was significantly lower under the canopies (40–80%) compared to open areas, which varied from 60 to 100% (ANOVA, P < 0.05), whereas amounts of ground vegetation cover, dead wood, and rocks did not differ (ANOVA, P > 0.05).
At the burned site, we found significantly higher values for both species richness and plant density inside the canopies of remnant trees for the plants in the smallest size class, i.e., the plants most likely to have established after the cyclones and fire. This pattern was further illustrated by the distribution of late-successional and vertebrate-dispersed Syzygium spp. (< 5 cm dbh), which showed on average that its density in the burned areas was four times higher inside the canopies than outside of them. Guevara et al. (1986) suggested that two factors are important in explaining the high abundance and diversity of woody plants under the canopies of isolated remnant trees: (1) higher seed deposition by frugivorous dispersers and (2) favorable microhabitats for the germination and establishment of late-successional species. Both mechanisms were probably operating in our study area. Our results showed that seed traps under the canopies of remnant trees in recently burned areas may receive 12 times more vertebrate-dispersed seeds than do traps outside of canopies. Also, seed traps under canopies may receive two to three times as many species of seeds as traps outside canopies. Shading was also higher under canopies, which likely favors seedling survival of late-successional plant species (Whitmore 1989). Overall, soil fertility was high in samples from all sites, but did not show any significant differences between plots inside and outside canopies.
For plants in larger size classes at the burned site in Study 1, no significant pattern was found except for medium-sized plants, which showed a marginally significant trend toward higher species richness outside canopies. Among larger plants, the probability is higher that they established before the cyclones and fire, so that their distribution was influenced more by predisturbance rather than postdisturbance dispersal processes.
At the crater site, no significant pattern at different distances from trees was found for any of the size classes, although there was a pronounced trend among the smallest plants toward a peak in both density and species richness at a distance of 5 m (at the margin or just outside of canopies). This pattern has still not been explained. Several factors probably contributed to a less pronounced contrast between conditions inside and outside of the canopies at the crater site. First, the absence of fire may explain, at least in part, why overall density and species richness are much higher (see Hjärpe et al. 2001). Second, because of the fourfold higher density of large trees (42 vs. 10/ha), the crater site probably experiences a much larger local passive seed rain, and regeneration is much less limited by vertebrate dispersal there than at the burned site (Hjärpe et al. 2001). Third, at the crater site the human impact is much higher; parts of the crater slope are used for agriculture, which may have influenced the distribution and abundance of the regenerating plant species in the remaining forest patches.
Our seed trap study lasted only two months (October and November), and most of the seeds found in the traps belonged to early-successional plants; few late-successional species were found. We lack local information on seasonal variation in fruit production, and it is possible that many late-successional species are dispersed at other times of the year. For example, Banack (1998) showed peaks for several late-successional species during May-September in American Samoa, based on an analysis of the diet of flying foxes (Pteropus spp.).
Two species of flying foxes (Pteropus tonganus and P. samoensis), starlings (Sturnus spp.), and several species of fruit pigeons (e.g., Ducula pacifica and Ptilinopus perousii) are potential dispersal vectors at both Tafua and Falealupo (Pierson et al. 1996). Flying foxes are known to have a very broad and generalized diet and forage for fruit on many canopy tree species in Samoa (Rainey et al. 1995, Pierson et al. 1996, Banack 1998). In areas with high flying fox activity, a pronounced multispecies composition of the seed rain under trees is expected (Banack 1998). In our limited study, however, the maximum number found in a single trap (less than 48 h of exposure) was three species, and the total number observed in all seed traps was 10 species.
Although the sources of the seeds of late-successional species are unknown, they most likely came from fruiting trees located in outside, less disturbed areas, because none of the remnant trees at the burned sites were on record as producing fruit. After both cyclones, it was possible to identify, based on satellite images, forest areas where the trees still retained many of their leaves and appeared to have been sheltered from the most destructive winds. These patches of forests were typically located on steep hillsides, in crevices, and in old volcano craters. Pierson et al. (1996) suggested that these areas functioned as refugia and constituted important sources for the colonization of the disturbed landscape for a large array of organisms. For at least the first few months after the cyclones and fires, such refugia may have been crucial sources for the dispersal of late-successional plant species and also important as feeding and support areas for vertebrate frugivores.
The results of this study are in agreement with the prediction that large remnant trees surviving in severely disturbed rain-forest areas represent biological legacies and serve as important nuclei for regeneration. However, we must still investigate the extent to which the observed patterns of high plant density and diversity under the canopies of remnant trees will be confined to the early-successional phases or continue to have a strong long-term influence on forest structure (e.g., Finegan 1996). Increased competition and thinning over time will probably change the composition and structure of the diverse local plant communities under remnant trees, so that these communities may, at least to some degree, be transient.
Holling et al. (1995) suggested that the crucial period after a destructive event (pest, fire, or storm), which they called the reorganization period, is often characterized by processes that restructure and reorganize species assemblages. During periods of this type, succession may be pushed along different pathways depending on the presence or absence of key factors in the initial phase (see also Nyström et al. 2000). In tropical forests that have been damaged by cyclones and fires, remnant trees and vertebrate frugivores may be viewed as key organisms in the reorganization phase. Their absence may force succession at burned sites along a different path, e.g., one that is dominated by wind-dispersed and passively dispersed plants, and may also lead to the risk of invasion by wind-dispersed, exotic, opportunistic species.
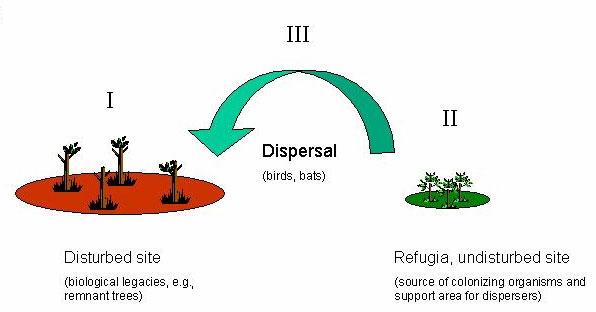
The design of forestry, conservation areas and other types of land use should, in general, be guided by the principle of maintaining the capacity of the ecosystem to buffer and reorganize after disturbances (e.g., Gunderson et al. 1995, Holling et al. 1995, Holling and Meffe 1996, Nyström et al. 2000). Based on this and previous studies (Elmqvist et al. 1994, Pierson et al. 1996, Hjärpe et al. 2001), we suggest that three factors represent essential components of the spatial resilience of tropical forest ecosystems and should be targeted for active management in tropical forests exposed to large-scale disturbances, particularly fire: (1) remnant trees, (2) refugia, and (3) vertebrate dispersers (Fig. 6).
|
Fig. 6. A conceptual model of proposed components of spatial resilience important for reorganization in tropical forests that have been disturbed by fires and cyclones.
|
Responses to this article are invited. If accepted for publication, your response will be hyperlinked to the article. To submit a comment, follow this link. To read comments already accepted, follow this link.
We would like to thank Ulu Taufaasisina and family in Tafua, Lilo Manuele and family in Falealupo, and Tautali and Vaasilifiti Moelagi for their help and guidance during the course of this study. Mats Arvidsson, James Atherton, Talie Foiga, Sifiti Tautali, and Nat Tuivavalagi also assisted in various ways. We also want to thank Dr. Art Whistler for his invaluable support in the identification of plant species. We also benefited from fruitful discussions with Johan Hjärpe, Henrik Hedenås, and Carl Folke. This study was financed in part by a NFR grant to T. Elmqvist and by Sida MFS grants to the other authors.
Banack, S. A. 1998. Diet selection and resource use by flying foxes (Genus Pteropus). 79:1949-1967.
Belsky, A. J., R. G. Amundson, J. M. Duxbury, S. J. Riha, A. R. Ali, and S. M. Mwonga. 1989. The effects of trees on their physical, chemical, and biological environments in a semi-arid savanna in Kenya. Journal of Applied Ecology 26:1005-1024.
Cox, P. A., and T. Elmqvist. 1991. Indigenous control of rain forest preserves: an alternative strategy for conservation. Ambio 20:317-321.
Cox, P. A., T. Elmqvist, E. D. Pierson, and W. E. Rainey. 1991. Flying foxes as strong interactors in South Pacific Island ecosystems: a conservation hypothesis. Conservation Biology 5:448-454.
Elmqvist, T., W. E. Rainey, E. D. Pierson, and P. A. Cox. 1994. Effects of tropical cyclones Ofa and Val on the structure of Samoan lowland rain forest. Biotropica 26:384-391.
Finegan, B. 1996. Pattern and process in neotropical secondary rain forests: the first 100 years of succession. Trends in Ecology and Evolution 11:119-123.
Foster, D. R., D. H. Knight, and J. F. Franklin. 1998. Landscape patterns and legacies resulting from large, infrequent forest disturbances. Ecosystems 1:497-510.
Franklin, J. F., and J. A. MacMahon. 2000. Messages from a mountain. Science 288:1183-1185.
Franklin, J. F., D. Lindenmayer, J. A. MacMahon, A. McKee, J. Magnuson, D. A. Perry, R. Waide, and D. Foster. 2000. Threads of continuity. Conservation Biology in Practice 1:8-16.
Guevara, S., S. E. Purata, and E. Van der Maarel. 1986. The role of remnant forest trees in tropical secondary succession. Vegetatio 66:77-84.
Guevara, S., J. Meave, P. Morenocasasola, and J. Laborde. 1992. Floristic composition and structure of vegetation under isolated trees in neotropical pastures. Journal of Vegetation Science 3:655-664.
Guevara, S., and J. Laborde. 1993. Monitoring seed dispersal at isolated remnant trees in tropical pastures: consequences for local species availability. Vegetatio 107/108:319-338.
Gunderson, L., C. S. Holling, and S. Light. 1995. Barriers and bridges to the renewal of ecosystems and institutions. Columbia University Press, New York, New York, USA.
Holling, C. S., D. W. Schindler, B. W. Walker, and J. Roughgarden. 1995. Biodiversity in the functioning of ecosystems: an ecological synthesis. Pages 44-83 in C. Perrings, K.-G. Mähler, C. Folke, C. S. Holling, and B.-O. Jansson, editors. Biodiversity loss: economic and ecological issues. Cambridge University Press, Cambridge, UK.
Holling, C. S., and G. K. Meffe. 1996. Command and control and the pathology of natural resource management. Conservation Biology 10:328-337.
Hjärpe, J., H. Hedenås, and T. Elmqvist. 2001. Post-cyclone recovery in a lowland rain forest in Samoa. Biotropica 33, in press.
Nyström, M., C. Folke, and F. Moberg. 2000. Coral reef disturbance and resilience in a human-dominated environment. Trends in Ecology and Evolution 15:413-417.
Pandaram, S. 1992. Tropical cyclone Val: tropical cyclone report 92/1. Fiji Meteorological Service, Suva, Fiji.
Pierson, E. D., T. Elmqvist, W. E. Rainey, and P. A. Cox. 1996. Effects of tropical cyclones on flying fox populations on the South Pacific islands of Samoa. Conservation Biology 10:438-451.
Rainey, W. E., E. D. Pierson, T. Elmqvist, and P. A. Cox. 1995. The role of the flying foxes (Pteropodidae) in oceanic island ecosystems of the Pacific. Zoological Symposium 67:47-62.
Robinson, G. R., and S. N. Handel. 1993. Forest restoration on a closed landfill: rapid addition of new species by bird dispersal. Conservation Biology 7:271-278.
Toh, I., M. Gillespie, and D. Lamb. 1999. The role of isolated trees in facilitating tree seedling recruitment at a degraded sub-tropical rainforest site. Restoration Ecology 7:288-297.
Turner, M. G., W. L. Baker, C. J. Peterson, and R. K. Peet. 1998. Factors influencing succession: lessons from large, infrequent natural disturbances. Ecosystems 1:511-523.
Whitmore, T. C. 1989. Canopy gaps and the two major groups of forest trees. Ecology 70:536-538.
Whistler, A. 1992. Vegetation of Samoa and Tonga. Pacific Science 46:159-178.
Wunderle, J. M. 1997. The role of animal seed dispersal in accelerating native forest regeneration on degraded tropical lands. Forest Ecology and Management 99:223-235.
Address of Correspondent:
Thomas Elmqvist
Department of Systems Ecology
Stockholm University
SE-106 91 Stockholm, Sweden
Phone: +46-8-161283
thomase@ecology.su.se
![]()