|
|
|
Copyright ©1997 by The Resilience Alliance*
Carpenter, S. R., and K. L. Cottingham. 1997. Resilience and restoration of lakes. Conservation Ecology [online]1(1): 2. Available from the Internet. URL: http://www.consecol.org/vol1/iss1/art2/A version of this article in which text, figures, tables, and appendices are separate files may be found by following this link.
Synthesis Resilience and Restoration of Lakes Stephen R. Carpenter1, Kathryn L. Cottingham2
1Center for Limnology, University of Wisconsin; 2National Center for Ecolological Analysis and Synthesis
- Abstract
- Introduction
- Resilience Mechanisms
- Pathological Dynamics
- Alternative States
- Resilience and Return Time
- Assessment of Resilience
- Significance of Biodiversity for Resilience
- Policies for Restoration
- Responses
- Literature Cited
KEY WORDS: ecological economics, ecosystem, eutrophication, lake, resilience, restoration, watershed.
Lakes provide humans with services that include water for irrigation, drinking, industry, and dilution of pollutants, hydroelectric power, transportation, recreation, fish, and esthetic enjoyment (Postel and Carpenter 1997). These services are impaired by exploitation of lakes and the lands of their catchments (Hasler 1947, Edmondson 1969, Harper 1992). Because human effects on lakes are growing, concern increases that lake ecosystem services are in jeopardy (Naiman et al. 1995). These concerns parallel those for the sustainability of services from many ecosystems and the biosphere itself (Arrow et al. 1995, Levin 1996).
Scientific studies of lake ecosystem processes have increased in spatial and disciplinary scope during the past century. Although many insights have been derived from the view of lakes as bounded systems, defined by the land-water interface, limnologists recognize that lakes must be understood in the landscape context of their catchments (Likens 1984, Wetzel 1990). Airborne pollutants and stratospheric ozone depletion connect lakes to perturbations of the global environment (D.W. Schindler et al. 1996). Changes in agriculture, riparian land use, forestry, fossil fuel consumption, and demand for ecosystem services link lakes to much larger social and economic systems (National Research Council 1993, Postel and Carpenter 1997).
Conceptual development has not kept pace with changes in the processes that alter and control lakes. Ecosystem ecology has a rich understanding of the physical, chemical, and biotic processes of lakes and their watersheds. There is a growing appreciation for the processes that explain heterogeneity among lakes in regional landscapes, regional effects of atmospheric deposition of contaminants in lakes, and the potential effects of global climate change on lakes (McKnight et al. 1996). However, we lack a conceptual framework for understanding the interactions of people and lakes. How do social and economic activities affect lakes? How do properties of lakes affect people's behavior toward lakes? What policies or institutions might sustain lakes and the services they provide? Such questions have barely been asked, let alone answered. Yet sustainable restoration of lakes must address the social and economic, as well as biotic and chemical, causes of lake degradation.
Concepts that integrate people and lakes must consider the processes that control normal and degraded lakes. Some relevant processes include climate, regional economic and social activity, watershed vegetation and land use, and internal components ranging from fish and macrophytes to phosphorus and humic staining. This paper describes the processes that control resilience of lakes, the pathology of degraded lakes, and lake restoration. Our goal is to synthesize published information and suggest some patterns and connections that contribute to an understanding of the interactions of lakes and people. The literature we review is drawn mostly from lake districts of North America and western Europe, where the most conspicuous human effects are associated with agriculture, urban development, industry, and recreation. However, our hypotheses about differences in control processes for normal and degraded lakes may apply to many of Earth's lake districts.
In this paper, a resilient system is one that tends to maintain a given state when subject to disturbance (Holling 1973, Ludwig et al. 1997). Lakes are routinely disturbed by many kinds of events. Inputs of solar radiation fluctuate from second to second. Pulses of chemical inputs occur with storms (at intervals of days to weeks) and annual snowmelt. Fluctuations in climate affect seasonal phenologies and budgets of heat, water, and nutrients from year to year (McKnight et al. 1996). Exceptionally large cohorts of fishes recruit every few years and restructure the food web for their lifetimes. Fires or other disturbances alter watershed vegetation at intervals of decades to centuries, thereby causing a pulse of nutrients to the lake and changing the water budget (through changes in evapotranspiration) as the vegetation regrows. Extreme changes in climate (with cycles of centuries) change hydrologic connections, water level, the shape of the lake, or even cause the lake to disappear for a period of time.
In the normal dynamics of lakes, ecosystem processes are maintained despite moderate and continuous disturbances originating in the lake, its watershed, and its airshed (Fig. 1 ). This resilience involves several mechanisms, which have different ecosystem components and distinctive spatial locations, spatial extents, and return times.
Fig. 1. Major interactions in the normal dynamics of lakes as a function of spatial extent (x-axis) and return time (y-axis). Boxes enclose 3 commonly-recognized subsystems: the watershed (largest box), lake (medium-size box), and pelagic zone (smallest box).
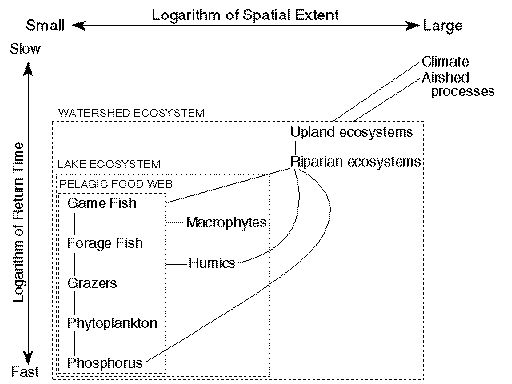
Riparian forests and grasslands delay or prevent nutrient transport from uplands to streams and lakes (Osborne and Kovacic 1993). Riparian forests are a source of fallen trees that can provide important fish habitat for decades (Maser and Sedell 1994, Christensen et al. 1996).
Wetlands function as vast sponges that delay the transport of water to downstream ecosystems and, thereby, reduce the risk of flooding (National Research Council 1992). Wetlands also modulate nutrient transport from uplands to streams and lakes (Johnston 1991). Wetlands are a major source of humic substances for lakes (Hemond 1990, Wetzel 1992). This complex of organic compounds stains lake water and affects ecosystem metabolism through several mechanisms (Jones 1992, Wetzel 1992). Of particular importance to this paper, humic staining suppresses the response of phytoplankton to pulses of nutrient input (Vollenweider 1976). This resilience mechanism involves shading, effects of humics on thermal structure of lakes, and changes in lake metabolism (Carpenter and Pace 1997).
Although phosphorus inputs and recycling establish the potential productivity of lakes (Schindler 1977), predation controls the allocation of phosphorus for production of fish, algal blooms, or other components of the pelagic food web (Carpenter and Kitchell 1993). In the normal dynamics of many lakes, large piscivorous game fishes are keystone predators that structure the food web below them (Kitchell and Carpenter 1993). Such lakes have large-bodied zooplankton grazers that effectively control phytoplankton (Carpenter et al. 1991). When pulses of phosphorus enter these lakes, the nutrient is transferred effectively to higher trophic levels and does not accumulate as algal biomass (Carpenter et al. 1996, D.E. Schindler et al. 1996).
Low or moderate rates of phosphorus input promote low rates of phosphorus recycling, through effects on the oxygen content of the water. Conditions of low-to-moderate productivity constrain respiration by bacteria, so that oxygen is not depleted from deeper waters during summer (Cornett and Rigler 1979). Oxygenated conditions decrease the rate of phosphorus recycling from sediments in many lakes (Caraco 1993). If production of the overlying water increases, deep waters can be deoxygenated and phosphorus recycling can increase, thereby further increasing production. Oxygenation of bottom waters prevents this positive feedback and confers resilience in moderately productive and unproductive lakes.
Submersed macrophytes of the littoral zone provide crucial habitat for attached algae, invertebrates, and fishes (Heck and Crowder 1991, Moss 1995). They also modify inputs to lakes from riparian or upstream ecosystems, store substantial amounts of nutrients, and are a source of dissolved organic compounds (Wetzel 1992). Oxygen production by macrophytes and attached algae can decrease the rate of phosphorus release from sediments, and high denitrification rates in littoral vegetation can decrease nitrogen availability (Wetzel 1992).
Collectively, these resilience mechanisms, operating at diverse scales, buffer lake ecosystems against fluctuating inputs. They maintain water quality, fish productivity, and the reliability of other ecosystem services provided to humans.
Measurements and scientific analyses of perturbations or resilience are always tied to particular scales of space and time (O'Neill et al. 1986). In the normal dynamics of lakes, perturbations are relatively brief in duration, but may be extensive in space. Examples are chemical or hydrologic fluctuations driven by weather, routine fluctuations of interacting populations, or fires that sweep through the watershed vegetation. Resilience mechanisms that tend to restore the normal dynamics involve longer or larger scales. Examples are food web dynamics that absorb nutrient pulses, wetlands that retain nutrients and release humic substances, or secondary succession of upland forests that stabilizes soils and retains nutrients. These resilience mechanisms can be destroyed by more extreme perturbations. Destruction of the normal resilience mechanisms is accompanied by the rise of new resilience mechanisms and qualitative changes in the ecosystem.
In normal lake dynamics, the dominant controlling processes are located in the watershed and the lake itself. In the pathological dynamics of lakes, the control shifts to processes with larger spatial extent (Fig. 2 ). Our use of "pathological" follows that of Leopold (1935): "Regarding society and land collectively as an organism, that organism has suddenly developed pathological symptoms, i.e., self-accelerating rather than self-compensating departures from normal functioning." Although the analogy of ecosystems to organisms now seems dated, many aspects of Leopold's essay seem current. Leopold recognized that both normal and degraded states of ecosystems could be self-sustaining. His concept of recuperative capacity ("capacity, when disturbed, to establish new and stable equilibria between soil, plants, and animals") is similar to the modern concept of resilience. Leopold also recognized that people and nature must be viewed as an integrated system to understand the self-sustaining properties of normal and degraded ecosystems.
Fig. 2. Major interactions in the pathological dynamics of lakes as a function of spatial extent (x-axis) and return time (y-axis). Italics denote control processes that are well developed in normal lakes, but are suppressed or absent in degraded lakes. Boxes correspond to those in Fig. 1.
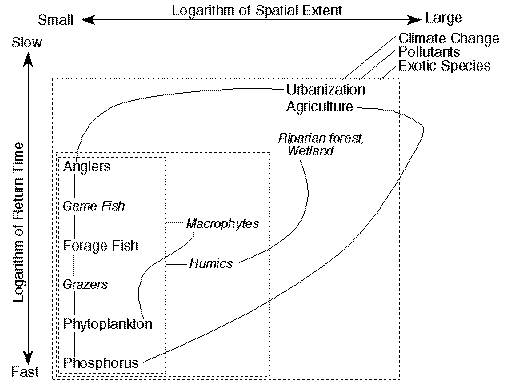
Degradation of lakes is a syndrome that involves breakdown of several resilience mechanisms and formation of several new ones (Fig. 2 ). The most common causes of lake degradation are pollutants from a variety of sources, especially agriculture (National Research Council 1992, 1993). The changes in land use that create and sustain pollutant flows are rooted in economic, demographic, and social changes that link lakes to large-scale human systems.
Eutrophication is probably the best understood type of lake degradation (Hasler 1947, Edmondson 1969, Harper 1992). Agriculture and urban development increase phosphorus transport to lakes. Losses of riparian vegetation and wetlands increase phosphorus flows. Humic inputs decline and humic constraints on phytoplankton become less effective. At the same time, game fish abundance is often reduced by overfishing, so planktivorous forage fishes and bottom-feeding fishes become more abundant. The large zooplanktonic grazers are reduced, and incoming phosphorus accumulates in phytoplankton biomass, especially in blue-green algae. Macrophyte beds decline because of reductions in water clarity and disturbance by bottom-feeding fishes. Loss of crucial habitat (macrophytes, wetlands, fallen trees) leads to further breakdown of the food web. The result is a lake with few piscivorous game fishes, abundant plankton- and bottom-feeding fishes, few large, herbivorous zooplankton, few macrophytes, dense algal blooms, and risks of anoxia and algal toxins. Toxic pollutants, species invasions, and species extirpations may interact with effects of phosphorus enrichment, habitat loss, and overfishing to exacerbate the degradation of lakes.
The dominant forces driving this degradation are regional to global in extent. Phosphorus pollution from manure and eroding cropland is a principal cause of lake degradation in the United States (National Research Council 1992). Farms import phosphorus, in the form of animal feeds to grow livestock, and as fertilizer to grow crops (National Research Council 1993). Some of the excess phosphorus drains into lakes to cause eutrophication (Daniel et al. 1994). Farmers' decisions about livestock densities and fertilizer applications are linked to regional or global markets for meat, dairy products, and grain.
Urban development, especially in wetland or riparian areas, is another principal cause of lake degradation (National Research Council 1992). Urban development is linked directly to human demography and regional economics. Likens (1992) identifies an important feedback in recreational lake districts called "leapfrog degradation." Riparian homeowners create lawns, remove shoreline vegetation, and harvest fish. Phosphorus inputs from fertilizers and septic systems promote algal growth, and losses of woody habitat and overfishing deplete fish stocks. The lake degrades, becoming less attractive for recreation. Consequently, development shifts to less disturbed lakes and the cycle is repeated.
Airborne pollutants and exotic species have become global controls of lake dynamics. Mercury, for example, is a widespread toxic pollutant derived from fossil fuels and other sources (Porcella et al. 1995). Under certain conditions, mercury is methylated and bioaccumulates through food chains (Driscoll et al. 1994). Organochlorine compounds are persistent toxic pollutants that affect many lakes. Although the more pernicious organochlorine compounds are no longer manufactured in the United States, significant amounts of some compounds still cycle within lakes and enter lakes from the atmosphere (Rasmussen et al. 1990, Stow et al. 1995). Acid precipitation has several negative impacts on lakes (Schindler et al. 1991), including a surprising interaction with stratospheric ozone depletion (D.W. Schindler et al. 1996). As lakes become more acid, humic concentrations decline and the water becomes more transparent to ultraviolet radiation, exacerbating the impacts on aquatic organisms of increased UV intensities, caused by declining ozone concentrations in the upper atmosphere.
Lakes, like islands, can be changed dramatically by species invasions. One spectacular example is the Laurentian Great Lakes, where invading sea lamprey (and overfishing) extirpated piscivorous lake trout, thereby allowing populations of another exotic species, the planktivorous alewife, to explode (Christie 1974). These exotics probably entered the Great Lakes through shipping canals. Fish stocking, ballast water, recreational boat traffic, and the aquarium trade have also transported exotics to new lakes. Some, but by no means all, invading species have powerful impacts on lakes, but our abilities to predict the effects of an invader in advance of the invasion are limited (Lodge 1993). Like the chemical changes in lakes, these biotic changes are driven by regional and global alterations of human activity and enterprise.
The pathological dynamics of lakes are also resilient. Ironically, this resilience can counter lake restoration efforts. In numerous cases, regulation of phosphorus in sewage effluents has been offset by increases in agricultural runoff or recycling from sediments (National Research Council 1992). Contaminant concentrations in fishes are stabilized by atmospheric inputs, recycling from sediments, and efficient trophic transfer (Stow et al. 1995). Restoration of piscivorous fishes has been countered by increases in angling (Kitchell 1992). To succeed, lake restoration programs must overcome the resilience of degraded lakes.
Alternative stable states (Ludwig et al. 1997) may be evident at several levels of watershed-lake systems. Watershed geochemistry, vegetation, and land use strongly affect lakes, and agricultural and urban development of watersheds usually shift lakes toward eutrophic states. Changes in watershed land use and fisheries management can cause abrupt and massive shifts in limnological conditions.
Many existent feedbacks act to stabilize lake trophic states of oligotrophy (low productivity, low nutrients, and clear water), dystrophy (low productivity and humic-stained water), and eutrophy (Wetzel 1983). These classically recognized states of lakes appear to be consistent with the concept of alternative stable states (Carpenter and Pace 1997). The interaction of humic inputs, phosphorus inputs, and planktonic metabolism can create alternative states of eutrophy and dystrophy under certain conditions. Rapid transitions between states can be caused by perturbations of chemical inputs or food web structure (Carpenter and Pace 1997).
Gradual changes in the planktivory rate can cause abrupt shifts in plankton community structure, grazing, algal biomass, and water clarity. Scheffer (1991) described this process using a model with two alternative states, turbid and clear. Dynamics of a cisco-zooplankton-phytoplankton food web in Lake Mendota, Wisconsin exhibit abrupt transitions that closely resemble alternative states (Rudstam et al. 1993). The model of Carpenter (1988) also exhibits turbid and clear states, but the dynamics follow an oscillation analogous to the spruce budworm system of Ludwig et al. (1997). In Carpenter's model, the oscillations are generated by a trophic cascade coupled to year classes of an age-structured piscivore population with density-dependent recruitment. Post et al. (1997) describe a case study of the cascading effects of largemouth bass year classes that corroborates the predictions of the model. The model also predicts that long-term fluctuations of phytoplankton should contain a cyclic component, with period roughly equal to the mean generation time of the dominant pelagic predator in the lake. Carpenter and Leavitt (1991) demonstrated such periodicities in a paleolimnological record.
The dynamics of shallow lakes provide the best limnological example of alternative states (Scheffer et al. 1993). Shallow eutrophic lakes exist in two states: turbid and dominated by phytoplankton, or clear and dominated by macrophytes (usually rooted aquatic plants, but sometimes attached macroalgae). The turbid state involves dense phytoplankton growth driven by nutrient recycling from sediments. Shading by phytoplankton blocks growth of attached plants. The plant-dominated state involves dense growths of attached plants that stabilize sediments (thereby slowing nutrient recycling) and that shelter phytoplankton grazers. The switch to the plant-dominated state can be triggered by a trophic cascade: piscivore stocking and/or planktivore removal to increase grazing and reduce phytoplankton (Jeppesen et al. 1990, van Donk and Gulati 1995, Moss et al. 1996). The switch to the turbid state can be triggered by water-level fluctuations (Blindow et al. 1993) or grazing of macrophytes by fish or birds (van Donk and Gulati 1995). Some lakes have switched states several times, with intervals of years to decades passing between transition events (Blindow et al. 1993).
Twenty-five years ago, resilience (Holling 1973) and return time (May 1974) were distinct concepts in ecology. Resilience is the capacity of a system to maintain certain structures and functions despite disturbance, and return time is the period required for a system to return to a particular configuration following disturbance. The return time to a stable equilibrium can be used to measure resilience of linear systems, or of nonlinear systems if the perturbations are small (Pimm 1984, DeAngelis 1992, Ludwig et al. 1997). In these situations, changes in return time can be used to measure changes in resilience.
In nonlinear ecosystems subject to large disturbances, however, losses of resilience are often associated with slow dynamics near an unstable point (Ludwig et al. 1997). In these situations, return times near stable points may be irrelevant. To address this issue, Holling (1973) and Ludwig et al. (1997) outline a broad view of resilience that depends on management objectives and options, the time scale of interest, and the type and magnitude of perturbations.
Although return rate near a stable point may miss important features of a system's resilience, empirical estimates of return rate may be useful for comparing responses of different systems to a given perturbation. Such measurements will usually be scale dependent, and there may be more than one possible explanation for an observed difference in return rate.
Return rate is related to nutrient turnover rate in diverse models of lake ecosystems (DeAngelis et al. 1989, Carpenter et al. 1992, DeAngelis 1992, Cottingham and Carpenter 1994). Phosphorus turnover rate is straightforward to measure and provides a means of testing hypotheses about lake ecosystem return rate (Cottingham and Carpenter 1994). These studies were conducted on lakes, not watersheds. Applications of this idea at watershed or regional scales have not yet been explored.
Analyses of experimental lakes have tested hypotheses about resilience and return rate. In lakes, shorter food chains have faster return rates (Carpenter et al. 1992), confirming a conjecture of Pimm and Lawton (1977). These fast return rates are due to rapid phosphorus dynamics of the smaller-bodied fishes that dominate the shorter food chains of lakes (Carpenter et al. 1992, Cottingham and Carpenter 1994). However, the shorter food chains are less capable of assimilating phosphorus pulses than are longer food chains (Carpenter et al. 1992, D.E. Schindler et al. 1996). In the short food chains, phosphorus pulses facilitate growth of grazing-resistant phytoplankton, and phosphorus accumulates as algae. Longer food chains contain larger bodied grazers that consume a wider range of phytoplankton species, and thereby transfer phosphorus to consumers more effectively than the smaller bodied grazers of shorter food chains (Carpenter and Kitchell 1993). Phosphorus pulses are absorbed by consumers or defecated to sediments in the longer food chains.
Recently, Ives (1995) proposed a method for estimating stochastic return time from noisy time series observations of ecosystems. He suggests that the ratio of variability in population growth rates to variability in population densities (estimated using regression models of a particular form) measures return rate for stochastic systems in a way analogous to return rate for deterministic systems. Ives' insight creates the opportunity to measure and compare stochastic return times for a wide variety of ecosystems, using actual time series data.
Ives' methods also suggest that longer food webs, dominated by large, piscivorous game fish, are more resilient to phosphorus inputs. Carpenter et al. (1996) measured grazer biomass and chlorophyll for four years in lakes with contrasting food webs that were enriched with phosphorus. Piscivore-dominated lakes contained largemouth bass, few planktivorous fishes, and large-bodied grazers, whereas planktivore-dominated lakes contained no piscivores and abundant planktivorous fishes and small-bodied grazers. The stochastic return rate was calculated for the total grazer biomass and the phytoplankton biomass. Ives' (1995) index of stochastic return rate was 0.89 in an unenriched piscivore-dominated lake, 0.51 in an enriched piscivore-dominated lake, and 0.10 in an enriched planktivore-dominated lake. These calculations show that phosphorus enrichment reduces stochastic return rate, and reduces it far more in the planktivore-dominated lake than in the piscivore-dominated lake.
In summary, resilience to phosphorus inputs in pelagic food webs depends on control of the phosphorus cycle by fishes. In piscivore-dominated lakes, the phosphorus return rate of the entire food web is slow because of the relatively slow phosphorus turnover rates of the fishes, which are a large phosphorus pool. However, by sharply reducing planktivory, the piscivores allow the development of a planktonic subsystem that efficiently converts phosphorus inputs to zooplankton biomass or sediment. Thus, phosphorus inputs do not accumulate as phytoplankton. If the piscivores are removed, planktivorous fishes dominate the food web and the phosphorus return rate of the entire food web increases, due to faster phosphorus turnover by the smaller bodied planktonic fishes. However, the efficient planktonic subsystem collapses, and is replaced by a subsystem in which phosphorus inputs accumulate as phytoplankton and symptoms of eutrophication are exacerbated.
Earth's lake districts contain ~100 million lakes >1 ha in area, and ~ 1 million lakes >1 km2 in area (Wetzel 1990). Lakes are often used as sentinel ecosystems, because they collect and integrate regional signals and preserve long-term information in their sediments. However, monitoring lakes at regional to continental scales is a significant challenge. Assessment of many lakes at the landscape scale, over sustained periods of time, requires relatively inexpensive and rapid assays. What indicators are appropriate for large-scale, long-term studies of the regional resilience of lake districts?
Resilience derives from partially redundant control processes that act at different scales to mitigate effects of perturbations. In normal lakes, key components are riparian vegetation, wetlands, game fish, and macrophytes. When these are intact, watershed-lake systems can withstand shocks at several scales, such as droughts, floods, forest fires, and recruitment variations. When these control mechanisms are broken down, control shifts to regional economic factors related to farm phosphorus budgets, development, and fishing. Perturbations translate into algal blooms and other symptoms of persistent eutrophication.
Our analysis suggests several potential indicators of a lake's capacity to maintain normal dynamics (Fig. 1 ). Livestock density in the watershed is a correlate of phosphorus imports (National Research Council 1993). Wetland area per unit lake area is an index of the landscape's capacity to hold water and export humic substances (Wetzel 1990). The proportion of the riparian zone occupied by forest and grassland indicates the potential attenuation of nutrient inputs (Osborne and Kovacic 1993). Lake color relates to humic content (Jones 1992). Slow-to-moderate piscivore growth rates are associated with strong piscivore control of planktivores (Kitchell et al. 1994). Grazer body size correlates with the capacity to suppress algal growth (Carpenter et al. 1991). Partial pressure of carbon dioxide in surface waters may be a sensitive indicator of ecosystem metabolism (Cole et al. 1994). Hypolimnetic oxygen depletion is a symptom of eutrophication and a driver of phosphorus recycling from sediments (Cornett and Rigler 1979, Caraco 1993). All of these indicators can be determined fairly inexpensively from agricultural data and land use records, remote sensing, and sampling to collect surface water, an oxygen profile, a zooplankton haul, fish size distribution, and fish scales for age determination. Calibration of these indicators using extant long-term records and broad regional surveys is a research priority.
The importance of biodiversity stems from the imbrication of control processes that maintain ecosystem resilience. Each control process may involve many species. The relative importance of a species for a given control process may vary from lake to lake or from time to time. Certain keystone species (Power et al. 1996) and "ecological engineers" (Jones and Lawton 1995) strongly influence ecosystem processes and resilience. Other species depend on the resilience mechanisms for their continued existence, but do not have obvious effects on resilience (Walker 1992). Thus, the association of biodiversity with resilience appears complex and variable.
Lake ecosystems can be configured in only a limited number of ways. Certain control processes are repeated in lake after lake, time after time. These regularities make it unnecessary to study each lake as if it were a new, unique system. Instead, we can look for general structures and processes that explain broad patterns.
Conservation of ecosystem structures is marvelously illustrated by Thingvallavatn, a deep, isolated, young lake carved by a glacier from a volcanic rift valley in Iceland (Campbell 1996). The food web of Thingvallavatn is dominated by arctic char. Over the past few thousand years, the chars have diversified from a common ancestor into four varieties: a planktivore, a small benthivore, a large benthivore, and a piscivore. The varieties differ substantially in maximum adult size, habitat use, mouth morphology, and diet. Thus, the ecosystem structure of Thingvallavatn closely resembles that of other lakes with more diverse fish assemblages. The resource polymorphisms of Thingvallavatn's arctic chars suggest that there are only certain limited ways in which lake ecosystems can be structured. This conservation of ecosystem structure resembles the regular clusters of adult body sizes described for terrestrial animal communities (Holling 1992, Holling et al. 1996).
Yet, how resilient is Thingvallavatn? A disturbance that affected arctic char could eliminate several ecosystem processes. Where one taxon controls an ecosystem process, species change and ecosystem change go hand in hand. Acidification of lakes, for example, eliminates a key group of bacteria, thereby blocking the cycle of nitrogen, one of the most important nutrients (Rudd et al. 1988). Would resilience be greater if several species were capable of performing each process? Experimental studies of more diverse lakes suggest that the answer is yes.
The role of species diversity in lake resilience is illustrated by experiments in which lakes were manipulated to various levels of toxic chemical stress or nutrient input. At low levels of toxic stress, changes in species composition are substantial, but changes in ecosystem process rates are negligible (Schindler 1990, Howarth 1991, Frost et al. 1995). Structural change at the species level stabilizes ecosystem process rates, an example of resilience called functional compensation by Frost et al. ( 1995). At more extreme levels of toxic chemical stress, functional compensation is not possible because too many species have been lost; consequently, ecosystem process rates change (Schindler 1990, Frost et al. 1995). Nutrient enrichment, in contrast, simultaneously changes both species composition and ecosystem process rates (Cottingham 1996). In this case, functional compensation allows ecosystem process rates to rapidly track shifts in availability of the limiting nutrient. Phytoplankton species t urnover rates are high at all enrichment levels. The availability of a large species pool may facilitate lake response to enrichment in the same way that high diversity facilitates grassland recovery from drought (Tilman 1996). Studies of both toxic str ess and nutrient enrichment show that we cannot predict which species will account for functional compensation or other responses to manipulation (Frost et al. 1995, Cottingham 1996). Ecosystem response to a given perturbation depends on only a fraction of the species pool, but the critical species are situation specific and can rarely be anticipated.
In summary, biodiversity confers resilience through compensatory shifts among species capable of performing key control processes. However, species number is not necessarily a good predictor of resilience. The link between biodiversity and resilience depends on the dominant ecosystem control processes, the complement of species capable of contributing to each process, and the susceptibility of these species to a particular ecosystem stress.
Breakdown of lake-watershed systems has been caused by policies that accelerate agricultural phosphorus flows, draining and development of wetlands, removal of riparian vegetation, overfishing, and spread of exotic species. The transformed, degraded state is also resilient. This resilience must be overcome to restore watersheds and lakes. But, according to the U.S. National Research Council (1992), "many so-called lake restoration projects really are only mitigation and management efforts to rid a lake, by whatever means, of some nuisance."
Reactive management, which responds to the symptom of the moment, but does not address systemic causes or long-term solutions, characterizes many water quality management programs (Soltero et al. 1992). Reactive management systems are subject to periodic upheavals, precipitated by crises that expose the inadequacies of current policy (Gunderson et al. 1995). In the United States, three cycles of water quality policy can be recognized (Carpenter et al. 1997). Public health concerns that arose in the course of federal water projects led to institutions for water treatment during the 1920s and 1930s. By about 1950, it was clear that lake eutrophication was widespread and unchecked. Management attention focused on point-source phosphorus controls, such as sewage treatment plants. As reductions of point-source phosphorus inputs became more common, it was evident by the 1970s that runoff from farms, construction sites, and urban areas was responsible for widespread degradation of lakes. We are still seeking institutional mechanisms capable of controlling runoff and restoring lakes degraded by pollution from diverse, disaggregated sources.
Sustainable restoration is not reactive; its goal "is to emulate a natural, self-regulating system that is integrated ecologically with the landscape in which it occurs" (National Research Council 1992). In other words, restoration requires shifting resilience mechanisms from those that maintain degraded systems (Fig. 2 ) to those that maintain more valuable systems (Fig. 1 ). Resilient restorations will tend to be self-sustaining. For lakes, resilient restorations require restoration of riparian, wetland, and macrophyte vegetation; reduction of phosphorus imports to farms; and reduced harvests of game fish. Ultimately, these changes are linked to social and economic processes at regional to continental scales.
The value of ecosystem services provided by lakes is substantial. Postel and Carpenter (1997) estimate that the total global value of freshwater ecosystem services is several trillion U.S. dollars per year. In the United States, the additional economic benefit of increasing lake water quality to meet acceptable standards for boating, fishing, and swimming is estimated at 31 to 55 billion U.S. dollars per year (Wadler et al. 1993; figures are adjusted to 1995 U.S. dollars). This calculation is an underestimate because it does not include benefits such as flood control, pollution dilution, reduced costs of purifying drinking water, and increased utility of cleaner water for irrigation and industry (Wadler et al. 1993). Degraded lakes represent the loss of substantial economic benefits.
In view of the economic benefits of clean lakes, it seems surprising that lakes continue to degrade and that restoration programs are few. The ecological causes of the problem are understood, and many useful technologies for lake restoration exist (National Research Council 1992). Why is restoration so difficult?
The fundamental problem of lake restoration is an economic mismatch: those who cause the problem do not benefit sufficiently from the remediation. On the other hand, the beneficiaries of lake restoration are not those who caused the degradation. The economic benefits of clean lakes need to be channeled in ways that create incentives for conservation of phosphorus on farms, restoration and maintenance of wetlands and riparian vegetation, and conservation of macrophytes and game fish. Thus far, the United States has not devised social and institutional mechanisms that achieve this fundamental goal.
Scientists can help in understanding this mismatch by shifting their analyses to an appropriately large reference frame. Aquatic scientists now study environments that are fundamentally different from those of the past. Although the basic biological and physical-chemical principles of limnology are unchanged, the dominant controls of lake ecosystems have shifted to much larger spatial scales. Ancestral local controls of lakes are much weakened and subordinated to new controls that involve regional trends in angling and fishing technology, economic and demographic forces that drive development, and agricultural markets that drive the phosphorus budgets of farms. The central question of applied limnology today is "How can we restore and sustain water quality, fisheries, and the other societal benefits of lakes in this new regime of regional to global control?" This question will not be answered by small-scale research, activism, or reactive management. It requires new, synthetic, fundamental studies of lakes and regulatory processes at multiple scales. The requisite understanding will come from a fusion of knowledge from natural and social sciences.
Resilience can stabilize valuable ecosystems or undesirable ones. Both valuable and degraded states of lakes can be self-sustaining. Agriculture and urbanization switch lakes from valuable to degraded states by changing the processes that control water quality and fisheries. However, lake degradation is not a necessary consequence of agriculture and urbanization. High-quality lakes can be maintained in developed landscapes. Restoration of lakes to the valuable state requires interventions that shift economic and ecological controls at several scales. Sustainable restoration will link the economic benefits of clean lakes to incentives for conservation.
Responses to this article are invited. If accepted for publication, your response will be hyperlinked to the article. To submit a comment, follow this link. To read comments already accepted, follow this link.
Acknowledgments
This paper was inspired by the Resilience Network meeting on Little St. Simon's Island, April 1996. We thank Terry Chapin, Tony Ives, Mike Pace, Mary Power, Daniel Schindler, and an anonymous referee for helpful comments on the manuscript. We are grateful for support from the Pew Foundation, the National Science Foundation, and the National Center for Ecological Analysis and Synthesis.
- Arrow, K., B. Bolin, R. Costanza, P. Dasgupta, C. Folke, C.S. Holling, B. Jansson, S. Levin, K. Maler, C. Perrings, and D. Pimentel. 1995. Economic growth, carrying capacity, and the environment. Science 268: 520-521.
- Blindow, I., G. Andersson, A. Hargeby, and S. Johansson. 1993. Long-term pattern of alternative stable states in two shallow eutrophic lakes. Freshwater Biology 30: 159-167.
- Campbell, D.G. 1996. Splendid isolation in Thingvallavatn. Natural History, June 1996: 48-55.
- Caraco, N.F. 1993. Disturbance of the phosphorus cycle: a case of indirect effects of human activity. Trends in Ecology and Evolution 8: 51-54.
- Carpenter, S.R. 1988. Transmission of variance through lake food webs. Pages 119-138 in S.R. Carpenter, editor. Complex interactions in lake communities. Springer-Verlag, New York, New York, USA.
- Carpenter, S.R., D. Bolgrien, R.C. Lathrop, C.A. Stow, T. Reed, and M.A. Wilson. 1997. Ecological and economic analysis of lake eutrophication by nonpoint pollution. Australian Journal of Ecology, in press.
- Carpenter, S.R., T.M. Frost, J.F. Kitchell, T.K. Kratz, D.W. Schindler, J. Shearer, W.G. Sprules, M.J. Vanni, and A.P. Zimmerman. 1991. Patterns of primary production and herbivory in 25 North American lake ecosystems. Pages 67-96 in J. Cole, S. Findlay, and G. Lovett , editors. Comparative analyses of ecosystems: patterns, mechanisms, and theories. Springer-Verlag, New York, New York, USA.
- Carpenter, S.R. and J.F. Kitchell, editors. 1993. The trophic cascade in lakes. Cambridge University Press, Cambridge, England.
- Carpenter, S.R., J.F. Kitchell, K.L. Cottingham, D.E. Schindler, D.L. Christensen, D.M. Post, and N. Voichick. 1996. Chlorophyll variability, nutrient input, and grazing: evidence from whole-lake experiments. Ecology 77: 725- 735.
- Carpenter, S.R., C.E. Kraft, R. Wright, X. He, P.A. Soranno, and J.R. Hodgson. 1992. Resilience and resistance of a lake phosphorus cycle before and after food web manipulation. American Naturalist 140: 781-798.
- Carpenter, S.R., and P.R. Leavitt. 1991. Temporal variation in a paleolimnological record arising from a trophic cascade. Ecology 72: 277-285.
- Carpenter, S.R., and M.L. Pace. 1997. Dystrophy and eutrophy in lake ecosystems: implications of fluctuating inputs. Oikos 78: 3-14.
- Christensen, D.L., B.R. Herwig, D.E. Schindler, and S.R. Carpenter. 1996. Impacts of lakeshore residential development on coarse woody debris in north temperate lakes. Ecological Applications 6: 1143-1149.
- Christie, W.J. 1974. Changes in the fish species compositions of the Great Lakes. Journal of the Fisheries Research Board of Canada 31: 827-854.
- Cole, J.J., N.F. Caraco, G.W. Kling, and T.K. Kratz. 1994. Carbon dioxide supersaturation in the surface waters of lakes. Science 265: 1568-1570.
- Cornett, R.J., and Rigler, F.H. 1979. Hypolimnetic oxygen deficits: their prediction and interpretation. Science 205: 580-581.
- Cottingham, K.L. 1996. Phytoplankton responses to whole-lake manipulations of nutrients and food webs. Dissertation. University of Wisconsin-Madison, Madison, Wisconsin, USA.
- Cottingham, K.L., and S.R. Carpenter. 1994. Predictive indices of ecosystem resilience in models of north temperate lakes. Ecology 75: 2127-2138.
- Daniel, T.C., A.N. Sharpley, D.R. Edwards, R. Wedepohl, and J.L. Lemunyon. 1994. Minimizing surface water eutrophication from agriculture by phosphorus management. Journal of Soil and Water Conservation 49: 30-38.
- DeAngelis, D.L. 1992. Dynamics of nutrient cycling and food webs. Chapman and Hall, New York, New York, USA.
- DeAngelis, D.L., S.M. Bartell, and A.L. Brenkert. 1989. Effects of nutrient recycling and food chain length on resilience. American Naturalist 134: 778-805.
- Driscoll, C.T., C. Yan, C.L. Schofield, R. Munson, and J. Holsapple. 1994. The mercury cycle and fish in Adirondack lakes. Environmental Science and Technology 28: 137-143.
- Edmondson, W.T. 1969. Eutrophication in North America. Pages 124-149 in Eutrophication: causes, consequences, correctives. National Academy Press, Washington, D.C., USA.
- Frost, T.M., S.R. Carpenter, A.R. Ives, and T.K. Kratz. 1995. Species compensation and complementarity in ecosystem function. Pages 224-239 in C. Jones and J. Lawton, editors. Linking species and ecosystems. Chapman and Hall , London, England.
- Gunderson, L.H., C.S. Holling, and S.S. Light, editors. 1995. Barriers and bridges to the renewal of ecosystems and institutions. Columbia University Press, New York, New York, USA.
- Harper, D. 1992. Eutrophication of freshwaters. Chapman and Hall, London, England.
- Hasler, A.D. 1947. Eutrophication of lakes by domestic drainage. Ecology 28: 383-395.
- Heck, K.L., and L.B. Crowder. 1991. Habitat structure and predator-prey interactions in vegetated aquatic systems. Pages 281-299 in S.S. Ball, E.D. McCoy, and H.R. Mushinsky, editors. Habitat structure: the physical arrangemen t of objects in space. Chapman and Hall, London, England.
- Hemond, H.F. 1990. Wetlands as the source of dissolved organic carbon to surface waters. Pages 301-313 in E.M. Perdue and E.T. Gjessing, editors. Organic acids in aquatic ecosystems. Wiley, New York, New York, USA.
- Holling, C.S. 1973. Resilience and stability of ecological systems. Annual Review of Ecology and Systematics 4: 1-23.
- --------------. 1992. Cross-scale morphology, geometry, and dynamics of ecosystems. Ecological Monographs 62: 447-502.
- Holling, C.S., G. Peterson, P. Marples, J. Sendzimir, K. Redford, L. Gunderson, and D. Lambert. 1996. Self-organization in ecosystems: lumpy geometries, periodicities and morphologies. Pages 346-384 in B.H. Walker and W.L. Steffen, editors. Global change and terrestrial ecosystems. Cambridge University Press, Cambridge, England.
- Howarth, R.W. 1991. Comparative responses of aquatic ecosystems to toxic chemical stress. Pages 169-195 in J. Cole, G. Lovett, and S. Findlay, editors. Comparative analyses of ecosystems. Springer-Verlag, New York, New Yor k, USA.
- Ives, A.R. 1995. Measuring resilience in stochastic systems. Ecological Monographs 65: 217-233.
- Jeppesen, E., J.P. Jensen, P. Kristensen, M. Sondergaard, E. Mortensen, O Sortkjaer, and K. Olrik. 1990. Fish manipulation as a lake restoration tool in shallow, eutrophic, temperate lakes. 2: Threshold levels, long-term stability, and c onclusions. Hydrobiologia 200/201: 219-227.
- Johnston, C.A. 1991. Sediment and nutrient retention by freshwater wetlands: effects on surface water quality. Critical Reviews in Environmental Control 21: 491-566.
- Jones, R.I. 1992. The influence of humic substances on lacustrine planktonic food chains. Hydrobiologia 229: 73-91.
- Jones, C., and J. Lawton, editors. 1995. Linking species and ecosystems. Chapman and Hall, London, England.
- Kitchell, J.F., editor. 1992. Food web management: a case study of Lake Mendota. Springer-Verlag, New York, New York, USA.
- Kitchell, J.F., and S.R. Carpenter. 1993. Variability in lake ecosystems: complex responses by the apical predator. Pages 111-124 in M. McDonnell and S. Pickett, editors. Humans as components of ecosystems. Springer-Verl ag, New York, New York, USA.
- Kitchell, J.F., E.A. Eby, X. He, D.E. Schindler, and R.M. Wright. 1994. Predator-prey dynamics in an ecosystem context. Journal of Fish Biology 45: 1-18.
- Leopold, A. 1935. Land pathology. Pages 212-217 in S.L. Flader and J.B. Callicott, editors. The river of the Mother of God and other essays by Aldo Leopold. 1991 edition. University of Wisconsin Press, Madison, Wisconsin, USA.
- Levin, S.A., editor. 1996. Forum: economic growth and environmental quality. Ecological Applications 6: 12-32.
- Likens, G.E. 1984. Beyond the shoreline: a watershed-ecosystem approach. Internationale Vereinigung fur Theoretische und Angewandte Limnology 22: 1-22.
- ------------. 1992. The ecosystem approach: its use and abuse. Ecology Institute, Oldendorf/Luhe, Germany.
- Lodge, D.M. 1993. Biological invasions: lessons for ecology. Trends in Ecology and Evolution 8: 133-137.
- Ludwig, D., B. Walker, and C.S. Holling. 1997. Sustainability, stability, and resilience. Conservation Ecology [online] 1 (1): 8 (http://www.consecol.org).
- Maser, C., and J.R. Sedell. 1994. From the forest to the sea: the ecology of wood in streams, rivers, estuaries, and oceans. St. Lucie Press, Delray Beach, Florida, USA.
- May, R.M. 1974. Stability and complexity in model ecosystems. Second edition. Princeton University Press, Princeton, New Jersey, USA.
- McKnight, D., D.F. Brakke, and P.J. Mulholland, editors. 1996. Freshwater ecosystems and climate change in North America. Limnology and Oceanography 41(6): 815-1149.
- Moss, B. 1995. The microwaterscape: a four-dimensional view of interactions among water chemistry, phytoplankton, periphyton, macrophytes, animals, and ourselves. Water Science and Technology 32: 105-116.
- Moss, B., J. Stansfield, K. Irvine, M. Perrow, and G. Phillips. 1996. Progressive restoration of a shallow lake: a 12-year experiment in isolation, sediment removal, and biomanipulation. Journal of Applied Ecology 33: 71-86 .
- Naiman, R.J., J.J. Magnuson, D.M. McKnight, and J.A. Stanford. 1995. The freshwater imperative. Island Press, Washington, D.C., USA.
- National Research Council. 1992. Restoration of aquatic ecosystems: science, technology, and public policy. National Academy Press, Washington, D.C., USA.
- ------------------. 1993. Soil and water quality: an agenda for agriculture. National Academy Press, Washington, D.C., USA.
- O'Neill, R.V., D.L. DeAngelis, J.B. Waide, and T.F.H. Allen. 1986. A hierarchical concept of ecosystems. Princeton University Press, Princeton, New Jersey, USA.
- Osborne, L.L., and D.A. Kovacic. 1993. Riparian vegetated buffer strips in water quality restoration and stream management. Freshwater Biology 29: 243-258.
- Pimm, S.L. 1984. The complexity and stability of ecosystems. Nature 307: 321-326.
- Pimm, S.L., and J.H. Lawton. 1977. Number of trophic levels in ecological communities. Nature 268: 329-331.
- Porcella, D.B., J.W. Huckabee, and B. Wheatley , editors. 1995. Mercury as a global pollutant. Kluwer, Amsterdam, The Netherlands.
- Post, D.M., S.R. Carpenter, D.L. Christensen, K.L. Cottingham, J.R. Hodgson, J.F. Kitchell, and D.E. Schindler. 1997. Seasonal effects of variable recruitment of a dominant piscivore on food web structure. Limnology and Oceanography, in press.
- Postel, S., and S.R. Carpenter. 1997. Freshwater ecosystem services. Pages 195-214 in G. Daily , editor. Nature's services. Island Press, Washington, D.C., USA.
- Power, M.E., D. Tilman, J.A. Estes, B.A. Menge, W.J. Bond, L.S. Mills, G. Daily, J.C. Castilla, J. Lubchenco, and R.T. Paine. 1996. Challenges in the quest for keystones. BioScience 46: 609-620.
- Rasmussen, J.B., R.B. Rowan, D.R.S. Lean, and J.H. Carey. 1990. Food chain structure in Ontario lakes determines PCB levels in lake trout (Salvelinus namaycush) and other pelagic fish. Canadian Journal of Fisheries and Aquatic S ciences 47: 2030-2038.
- Rudd, J.W.M., C.A. Kelly, D.W. Schindler, and M.A. Turner. 1988. Disruption of the nitrogen cycle in acidified lakes. Science 240: 1515-1518.
- Rudstam, L.G., R.C. Lathrop, and S.R. Carpenter. 1993. The rise and fall of a dominant planktivore: direct and indirect effects on zooplankton. Ecology 74: 303-319.
- Scheffer, M. 1991. Fish and nutrients interplay determines algal biomass: a minimal model. Oikos 62: 271-282.
- Scheffer, M., S.H. Hosper, M.-L. Meijer, B. Moss, and E. Jeppesen. 1993. Alternative equilibria in shallow lakes. Trends in Ecology and Evolution 8: 275-279.
- Schindler, D.E., S.R. Carpenter, K.L. Cottingham, X. He, J.R. Hodgson, J.F. Kitchell, and P.A. Soranno. 1996. Food web structure and littoral coupling to pelagic trophic cascades. Pages 96-108 in G.A. Polis and K.O. Winemiller, edi tors. Food webs: integration of pattern and dynamics. Chapman and Hall, New York, New York, USA.
- Schindler, D.W. 1977. The evolution of phosphorus limitation in lakes: natural mechanisms compensate for deficiencies of nitrogen and carbon in eutrophied lakes. Science 195: 260-262.
- ----------------. 1990. Experimental perturbations of whole lakes as tests of hypotheses concerning ecosystem structure and function. Oikos 57: 25-41.
- Schindler, D.W., P.J. Curtis, B.R. Parker, and M.P. Stainton. 1996. Consequences of climate warming and lake acidification for UV-B penetration in North American boreal lakes. Nature 379: 705-708.
- Schindler, D.W., T.M. Frost, K.H. Mills, P.S.S. Chang, I.J. Davies, D. Findlay, D.F. Malley, J.A. Shearer, M.A. Turner, P.J. Garrison, C.J. Watras, K. Webster, J.M. Gunn, P.L. Brezonik, and W.A. Swenson. 1991. Comparisons betweeen experimen tally and atmospherically acidified lakes. Proceedings of the Royal Society of Edinburgh 97B: 193-226.
- Soltero, R.A., L.R. Singleton, and C.R. Patmont. 1992. The changing Spokane River watershed: actions to improve and maintain water quality. Pages 458-478 in R.J. Naiman, editor. Watershed management. Springer-Verlag, New Y ork, New York, USA.
- Stow, C.A., S.R. Carpenter, C.P. Madenjian, L.A. Eby, and L.J. Jackson. 1995. Fisheries management to reduce contaminant consumption. BioScience 46: 752-758.
- Tilman, D. 1996. Biodiversity: population vs. ecosystem stability. Ecology 77: 350-363.
- van Donk, E., and R.D. Gulati. 1995. Transition of a lake to turbid state six years after biomanipulation: mechanisms and pathways. Water Science and Technology 32: 197-206.
- Vollenweider, R.A. 1976. Advances in defining critical loading levels for phosphorus in lake eutrophication. Memorie dell'Istituto Italiano di Idrobiologia 33: 53-83.
- Wadler, R., J.C. Landman, and D.M. Cameron. 1993. The Clean Water Act 20 years later. Island Press, Washington, D.C., USA.
- Walker, B.H. 1992. Biological diversity and ecological redundancy. Conservation Biology 6: 18-23.
- Wetzel, R.G. 1983. Limnology. W.B. Saunders, Philadelphia, Pennsylvania, USA.
- -----------. 1990. Land-water interfaces: metabolic and limnological regulators. Internationale Vereinigung fur Theoretische und Angewandte Limnology 24: 6-24.
- -----------. 1992. Gradient-dominated ecosystems: sources and regulatory functions of dissolved organic matter in freshwater ecosystems. Hydrobiologia 229: 181-198.
- Blindow, I., G. Andersson, A. Hargeby, and S. Johansson. 1993. Long-term pattern of alternative stable states in two shallow eutrophic lakes. Freshwater Biology 30: 159-167.
Address of Correspondent:
Stephen R. Carpenter
Center for Limnology
680 North Park Street
University of Wisconsin
Madison, Wisconsin 53706 USA
phone: 608-262-8690
fax: 608-265-2340
srcarpen@facstaff.wisc.edu
*The copyright to this article passed from the Ecological Society of America to the Resilience Alliance on 1 January 2000.
![]()